
Biological Cybernetics (2021) 115:539–562
https://doi.org/10.1007/s00422-021-00899-1
ORIGINAL ARTICLE
Mapping input noise to escape noise in integrate-and-fire neurons: a
level-crossing approach
Tilo Schwalger1,2
Received: 17 July 2021 / Accepted: 27 September 2021 / Published online: 19 October 2021
© The Author(s) 2021
Abstract
Noise in spiking neurons is commonly modeled by a noisy input current or by generating output spikes stochastically with a
voltage-dependent hazard rate (“escape noise”). While input noise lends itself to modeling biophysical noise processes, the
phenomenological escape noise is mathematically more tractable. Using the level-crossing theory for differentiable Gaussian
processes, we derive an approximate mapping between colored input noise and escape noise in leaky integrate-and-fire
neurons. This mapping requires the first-passage-time (FPT) density of an overdamped Brownian particle driven by colored
noise with respect to an arbitrarily moving boundary. Starting from the Wiener–Rice series for the FPT density, we apply the
second-order decoupling approximation of Stratonovich to the case of moving boundaries and derive a simplified hazard-rate
representation that is local in time and numerically efficient. This simplification requires the calculation of the non-stationary
auto-correlation function of the level-crossing process: For exponentially correlated input noise (Ornstein–Uhlenbeck process),
we obtain an exact formula for the zero-lag auto-correlation as a function of noise parameters, mean membrane potential and
its speed, as well as an exponential approximation of the full auto-correlation function. The theory well predicts the FPT and
interspike interval densities as well as the population activities obtained from simulations with colored input noise and time-
dependent stimulus or boundary. The agreement with simulations is strongly enhanced across the sub- and suprathreshold
firing regime compared to a first-order decoupling approximation that neglects correlations between level crossings. The
second-order approximation also improves upon a previously proposed theory in the subthreshold regime. Depending on a
simplicity-accuracy trade-off, all considered approximations represent useful mappings from colored input noise to escape
noise, enabling progress in the theory of neuronal population dynamics.
Keywords Integrate-and-fire neuron ·Interspike interval density ·First-passage-time density ·Colored noise ·Escape noise ·
Hazard rate ·Threshold-crossing statistics ·Neuronal population dynamics
1 Introduction
Neurons in the brain must operate under highly non-
stationary conditions. In fact, most behaviorally relevant
sensory stimuli as well as internal signals are rarely con-
stant in time but may change rapidly. In the presence of
noise, such dynamic stimuli can be reliably encoded in the
time-dependent population activity of a large population of
Communicated by Moritz Helias.
BTilo Schwalger
1Institute of Mathematics, Technical University Berlin, 10623
Berlin, Germany
2Bernstein Center for Computational Neuroscience Berlin,
10115 Berlin, Germany
spiking neurons (Gerstner et al. 2014). The time-dependent
population activity also provides a concise coarse-grained
description of the collective dynamics of interacting spik-
ing neurons. Therefore, theories that predict the population
activity in response to a time-dependent signal have been
of fundamental interest in theoretical neuroscience (Knight
1972; Gerstner 2000; Augustin et al. 2017; Schwalger et al.
2017).
The population activity of noisy spiking neurons can be
mathematically described by population density equations
(Nykamp and Tranchina 2000; Chizhov 2017). The form
of the population density equation depends on the noise
model. Two popular ways to model neuronal noise consist
of modeling noise either in the input or in the output of the
neuron (Gerstner et al. 2014). In the first model class (input
noise), noise enters the dynamical equations of the membrane
123

540 Biological Cybernetics (2021) 115:539–562
potential, currents or conductances leading to stochastic dif-
ferential equations. If the noise is Gaussian white noise, the
subthreshold dynamics becomes a diffusion process and the
input noise is also called diffusive noise (Gerstner 2000).
The corresponding population density equation is a Fokker–
Planck equation and the population activity can be obtained
as the probability flux across the threshold (Abbott and
van Vreeswijk 1993; Brunel and Hakim 1999; Fourcaud
and Brunel 2002; Nykamp and Tranchina 2000; Richard-
son 2008; Augustin et al. 2017). Models based on diffusive
noise naturally appear as the result of modeling biophysical
processes such as synaptic shot-noise or ion channel noise.
In particular, a frequently considered source of noise is back-
ground synaptic input modeled as external Poisson processes
(Brunel 2000; Potjans and Diesmann 2014). The fluctuating
part of this external shot noise leads, via a diffusion approxi-
mation (Gerstner et al. 2014), to Gaussian white noise driving
the synaptic input current or conductance. Besides its bio-
physical interpretability, input noise has the advantage that it
permits modeling both temporal (Fourcaud and Brunel 2002;
Schwalger et al. 2015) and spatial (Lindner et al. 2005) corre-
lations of synaptic inputs and enables mean-field theories for
recurrent networks of sparsely connected integrate-and-fire
neurons (Brunel and Hakim 1999; Brunel 2000).
In the second model class (called output noise or escape
noise (Gerstner 2000)), the dynamical equations for the state
variables are deterministic, while spikes (“output”) are gen-
erated stochastically through a hazard rate or conditional
intensity (Gerstner 2000; Paninski 2004; Truccolo et al.
2005; Pillow et al. 2008; Pillow and Latham 2008; Naud
and Gerstner 2012;Breaetal.2013; Galves and Löcherbach
2016; Gerhard et al. 2017; Raad et al. 2020). This hazard
rate depends on the state variables via a link function. For
example, it may be given as ˆ
λ(t)=Ψ(u(t), ˆ
t(t)), where
u(t)is the membrane potential and ˆ
t(t)is the last spike
time of the neuron at time t. If the neuron model is a
non-homogeneous renewal or quasi-renewal (Naud and Ger-
stner 2012) process, the corresponding population density
equation is a renewal integral equation or, equivalently, a
refractory density equation (Gerstner et al. 2014; Gerstner
2000; Naud and Gerstner 2012; Chizhov and Graham 2007,
2008; Dumont et al. 2016; Schwalger and Chizhov 2019).
Although output noise is of phenomenological nature without
a quantitative link to biophysical mechanisms, it has several
advantages (Schwalger and Chizhov 2019) owing to its sim-
pler mathematical tractability: First, the refractory density
or integral equation admits an extension to finite numbers
of neurons (Schwalger et al. 2017; Schwalger and Chizhov
2019; Schmutz et al. 2020,2021). This extension allows to
account for finite-size fluctuations of the population activ-
ity at the mesoscopic scale. Second, models with output
noise provide analytical expressions for the likelihood func-
tion, and thus, model parameters can be efficiently fitted
to experimental data of single neuron recordings (Panin-
ski 2004; Truccolo et al. 2005; Pillow et al. 2008; Gerhard
et al. 2017;Mensietal.2012; Pozzorini et al. 2015; Teeter
et al. 2018). And third, the state space for models with
output noise remains approximately one-dimensional even
for multi-dimensional conductance-based neuron models
(Chizhov and Graham 2007). The one-dimensional descrip-
tion permits highly efficient numerical solutions, in contrast
to Fokker–Planck equations (Apfaltrer et al. 2006), which
become intractable and computationally inefficient for sev-
eral state variables.
In view of the wide use of biologically interpretable input
noise and the mathematical advantages of output noise, an
intriguing question is whether input noise can be approxi-
mately mapped to output noise, so as to take full advantage
of both noise models. Mathematically, such a map requires
the specification of the hazard rate ˆ
λ(t)in terms of a link
function Ψ, which depends on some dynamical variables
and defines the escape-noise model. Unfortunately, a stan-
dard method to derive such a link function does not exist.
To see this, let us consider the example of non-homogeneous
renewal processes as a popular class of neuron models. In
these models, the probability density P(t|ˆ
t)to fire the next
spike at time tgiven a spike at time ˆ
t,ˆ
t<t, does not depend
on the state of the model before time ˆ
t, i.e., the memory
of renewal neurons only reaches back to its last spike. An
important example of non-homogeneous renewal models in
neuroscience are one-dimensional integrate-and-fire neurons
driven by white input noise (Gerstner et al. 2014). For this
model class one can formally construct the hazard rate via
the formula λ(t|ˆ
t)=P(t|ˆ
t)/1−t
ˆ
tP(s|ˆ
t)ds(Gerstner
et al. 2014). However, there are two obstacles: first, in order
to apply this formula, the “interspike interval (ISI) density”
P(t|ˆ
t)would be needed in analytical form for arbitrary, time-
dependent input currents {I(t)}t∈(ˆ
t,t)that occurred since the
last spike. However, the calculation of the ISI density for
time-dependent inputs is equivalent to a first-passage-time
(FPT) problem with time-dependent parameters or boundary.
The solution of this FPT problem requires the solution of the
Fokker–Planck equation with moving absorbing boundary,
which is known to be a hard theoretical problem (Bulsara
et al. 1996; Schindler et al. 2004; Lindner 2004b). Second,
even if one succeeds to derive an approximate formula for
the hazard rate λ(t|ˆ
t), it is still challenging to represent the
hazard rate in the form of a link function Ψu(t), {z(t)},ˆ
t
that depends on some voltage-like variable u(t),thelast
spike time ˆ
tand possibly further dynamical variables {z(t)}
locally in time (as opposed to a “non-local” functional of
{u(t), z(t)}t∈(ˆ
t,t)).
Several theoretical studies have suggested approximate
local hazard rates for leaky integrate-and fire (LIF) models
driven by white (Plesser and Gerstner 2000; Herrmann and
Gerstner 2001; Chizhov and Graham 2007) or exponentially
123

Biological Cybernetics (2021) 115:539–562 541
correlated (Chizhov and Graham 2008) Gaussian noise, or
quasi-static (frozen) noise (Goedeke and Diesmann 2008).
In this paper, we explore an alternative approach to the haz-
ard rate and the first-passage-time density based on the theory
of level crossings (Verechtchaguina et al. 2006). In Sect. 2,
we introduce the LIF model with time-dependent driving
and constant threshold and map this process an equivalent
model with constant input and moving barrier. In Sect. 3,
we consider the level crossing statistics with respect to this
moving barrier and use the Wiener–Rice series and approx-
imations thereof to provide formal expressions for the FPT
density. These expressions form the starting point for deriv-
ing approximate hazard rates that are local in time. This
derivation reveals some unexpected results concerning the
correlations of level-crossings of Gaussian processes at small
time lags (Sect. 3.4). Then, we turn to the LIF model and the
problem of mapping input noise to escape noise (Sect. 4)
and apply this map to predict the time-dependent population
activity of LIF neurons with colored input noise (Sect. 5).
Each of the Sects. 3,4and 5closes with a comparison of the
level-crossing theory with simulations and a previous the-
ory by Chizhov and Graham (2008). Detailed derivations are
provided in Appendix.
2 Leaky integrate-and-fire models and the
associated first-passage-time problem
As a spiking neuron model with input noise, we consider a
leaky integrate-and-fire model driven by synaptically filtered
(“colored”) noise (Schwalger and Schimansky-Geier 2008;
Gerstner et al. 2014; Schuecker et al. 2015). In this model,
spikes are emitted whenever the membrane potential V(t)
reaches a threshold VT. The subthreshold dynamics for V<
VTcan be written as
τm˙
V=−V+μ(t)+η(t), (1a)
τs˙η=−η+2τsσηξ(t), (1b)
where τmis the membrane time constant and μ(t)=Vrest +
RI(t)is the mean neuronal drive consisting of a constant
resting potential Vrest and a time-dependent input current
I(t)(Rdenotes the membrane resistance). Furthermore, η(t)
is a colored noise modeled as a one-dimensional Ornstein–
Uhlenbeck process with correlation time τsand variance σ2
η,
and ξ(t)is a zero-mean Gaussian white noise with auto-
correlation function ξ(t)ξ(t)=δ(t−t). The colored
noise captures the effect of various intrinsic and extrinsic
noise sources, such as fluctuations of synaptic background
activity in vivo (shot noise due to random spike arrival from
background neurons). After threshold crossing and spike
emission, V(t)is reset to a reset potential VR,VR<VT, and
the subthreshold dynamics Eq. (1) resumes after an absolute
refractory period of length tref following the reset.
We are seeking a corresponding spiking neuron model
with escape-noise (Gerstner 2000) given by a hazard rate
(conditional intensity) of the form Ψu(t), ˙u(t), {zi(t)},t−
ˆ
t. Here, u(t)is a membrane-potential variable that obeys
the noiseless membrane dynamics of the LIF model between
spikes:
τm˙u=−u+μ(t). (2a)
Furthermore, we allow an explicit dependence on the speed
of the membrane potential ˙u(t)(in accordance with previous
studies (Plesser and Gerstner 2000; Herrmann and Gerstner
2001; Chizhov and Graham 2007; Goedeke and Diesmann
2008)), the time since the last spike t−ˆ
t, and possibly fur-
ther auxiliary variables {zi}whose dynamics between spikes
is given by ordinary differential equations. Given these vari-
ables at time t, a spike is fired independently in the next time
step with probability
Pr spike in (t,t+dt)|u(t), ˙u(t), {zi(t)},t−ˆ
t
=Ψu(t), ˙u(t), {zi(t)},t−ˆ
tdt(2b)
where dtis a small step size. This probabilistic firing rule
is the counterpart of the firing rule with a hard threshold
in the LIF model with input noise. After a spike, u(t)is
reset to VRand the auxiliary variables {zi}are also reset to
some suitable fixed reset value. During an absolute refractory
period of length tref, the variables are clamped to their reset
values and the hazard rate is set to zero. Because all memory
is erased upon resetting, the escape-noise model is a non-
homogeneous renewal process .
The main goal is to map the model with colored input
noise, Eq. (1) to the model with escape noise, Eq. (2). Strictly
speaking, mapping the two models is an ill-posed problem
because the model with input noise is a non-renewal process,
whereas the escape-noise model is a (non-homogeneous)
renewal process. In fact, the temporal correlations of the col-
ored noise in Eq. (1) introduces memory that is not erased
upon spiking. This memory leads to correlations between
interspike intervals (ISIs) (Lindner 2004a; Schwalger and
Schimansky-Geier 2008; Schwalger et al. 2015). However,
if the correlation time τsof the colored noise is much smaller
than the mean interspike interval, these correlations will be
small and the model with input noise can be regarded as
approximately renewal. In this case, it is sufficient to match
the ISI densities of the two models in order to obtain an
approximate mapping. Therefore, our goal of mapping the
two models can be phrased more modestly as follows: Can
we find a link function Ψof the escape-noise model such
that for an arbitrary given stimulus μ(t)the time-dependent
ISI densities P(t|ˆ
t)of the two models approximately match
123
542 Biological Cybernetics (2021) 115:539–562
for all tand ˆ
t<t? We emphasize that this definition of the
mapping rests on the assumption of sufficiently small cor-
relation times of the colored input noise. Biologically, this
assumption seems to be reasonable given that typical time
scales of excitatory and inhibitory postsynaptic currents are
often only on the order of a few milliseconds (Gerstner et al.
2014).
To derive the link function Ψthat maps input to output
noise, one needs to solve a first-passage-time (FPT) prob-
lem: As mentioned in Introduction, the hazard rate can be
obtained from the ISI density of the model with input noise,
Eq. (1). In this model, the interspike interval is determined
by the “first-passage time” that is needed for the membrane
potential to travel from the reset potential to the threshold.
Thus, the ISI density P(t|ˆ
t)is equivalent to the FPT den-
sity (apart from a time shift due to the deterministic absolute
refractory period). To compute the FPT density, one needs
to choose suitable initial conditions for the colored noise
η(t). The ISI starting at the last spike time ˆ
tis composed
of the initial absolute refractory period of length tref and the
stochastic FPT t∗. We thus need the initial value η(ˆ
t+tref)
of the noise at the starting time ˆ
t+tref of the stochastic
motion. At the firing time ˆ
t, the distribution of the noise
pfire(η, ˆ
t)is biased toward positive values of η(Lindner
2004a; Schwalger and Schimansky-Geier 2008; Schwalger
2013; Schwalger et al. 2015), in contrast to the stationary dis-
tribution pst(η) of the Ornstein–Uhlenbeck noise, which has
zero mean. During the absolute refractory period, the noise
distribution relaxes toward the stationary distribution. Even
though the noise at time ˆ
t+tref may not be fully stationary
yet, it is reasonable to assume stationary initial conditions,
where η(ˆ
t+tref)∼N(0,σ2
η)is drawn from a normal dis-
tribution with variance σ2
η. This initial condition is justified
because the noise correlation time τshas been assumed to
be much smaller than the mean ISI; hence, we do not expect
that the precise shape of the initial noise distribution has a
significant effect on the FPT density.
Because in the following we focus on the FPT starting at
ˆ
t+tref, we will conveniently choose the time origin such that
ˆ
t+tref =0. Furthermore, since we are only interested in
the first threshold crossing after time t=0, we can omit the
voltage resetting for t>0 without changing the FPT statis-
tics. The resulting non-resetting process ˆ
V(t)is the freely
evolving solution of Eq. (1) without reset and with initial
conditions ˆ
V(0)=VR,η(0)∼N(0,σ2
η)(Fig. 1a). This
non-resetting process will be useful for the level-crossing
approach below.
For mathematical convenience, we will now reformulate
the FPT problem in terms of a time-homogeneous process
x(t)and a moving boundary b(t), so as to eliminate the
time-dependent parameter μ(t)in Eq. (1) (Fig. 1b). This is
achieved by subtracting the mean non-resetting membrane
potential ˆ
V(t)=u(t):
(a)
(b)
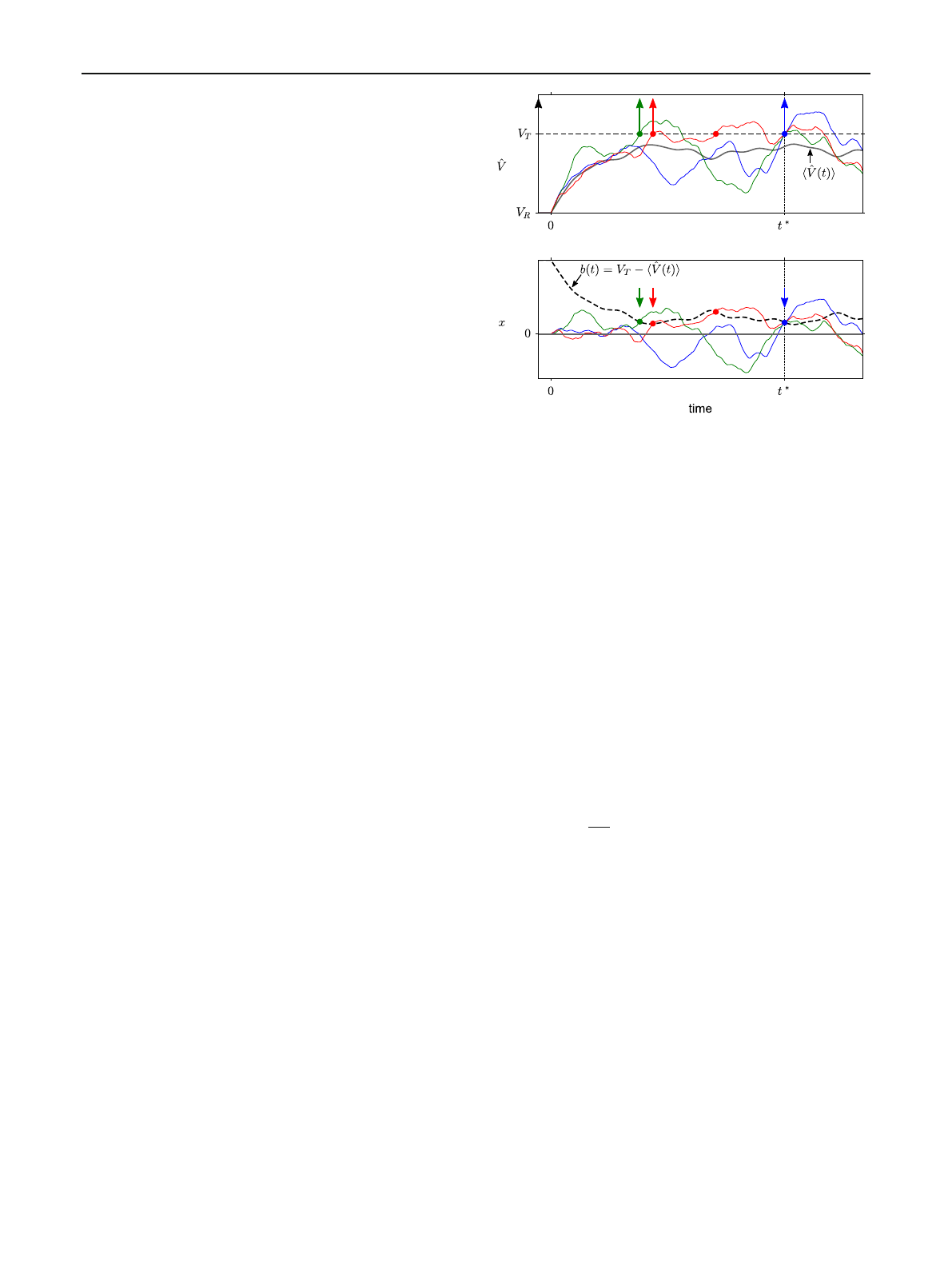
Fig. 1 First-passage time of an integrate-and-fire neuron model and
an equivalent model with moving boundary. aAt time t=0, differ-
ent realizations of the non-resetting membrane potential ˆ
V(t)(colored
thin lines) are released from the reset potential VR. The non-resetting
membrane potential follows a Gaussian process with time-dependent
mean ˆ
V(t)(gray thick line). Shown are three realizations (green, red,
blue lines) that have an identical threshold crossing at time t=t∗
(blue circle), which is not necessarily the first crossing (indicated by an
arrow). bTransformation to an equivalent time-homogeneous process
x(t)with moving boundary b(t), in which the positions of thresh-
old crossings are preserved. Parameters: τs=4 ms, τm=10 ms,
σV:=σx(∞)=0.25(VT−VR)
x(t)=ˆ
V(t)−u(t)(3)
b(t)=VT−u(t), (4)
where u(t)is given by Eq. (2a) with initial condition u(0)=
VR. Furthermore, setting y=η/τm,γ=1/τm,τy=τsand
D=τsσ2
η/τ2
m, we find the Langevin equation
˙x=−γx+y(5a)
τy˙y=−y+√2Dξ(t)(5b)
with initial conditions
x(0)=0,y(0)∼N(0,σ2
y)(6)
The dynamics of x(t)can be interpreted as an overdamped
motion of Brownian particle in a parabolic potential sub-
ject to a colored noise y(t)(Ornstein–Uhlenbeck process).
Here, Dand τyare intensity and the correlation time of the
noise, respectively, and γis the friction coefficient. As before,
ξ(t)is a zero-mean Gaussian white noise. At time t=0,
the random initial condition for the colored noise ycorre-
sponds to a stationary Gaussian distribution with mean zero
and variance σ2
y=D/τy. By construction, the domain of
the particle is bounded from above by the time-dependent
123

Biological Cybernetics (2021) 115:539–562 543
boundary b(t), where b(0)>0 and b(t)is a differentiable
function of time. The FPT t∗is defined as the time when
x(t)exits the domain, i.e., when it reaches the boundary,
for the first time. The FPT density will be denoted by P(t),
i.e., P(t)dt=Prob(t∗∈[t,t+dt)) for an infinitesimal
time interval of length dt. We emphasize again that the FPT
density of the Brownian particle x(t)with moving boundary
b(t)is the same as the FPT density of the membrane potential
V(t)with respect to the constant threshold VT.
Beyond neuroscience, the escape of the doubly low-
pass filtered process, Eq. (5), from a domain with moving
boundary b(t)may serve as a simple archetypal model for
non-stationary FPT problems. One prominent example is
reaction times of bimolecular chemical reactions (Hänggi
et al. 1990). If x(t)is interpreted as a reaction coordinate and
the domain x<b(t)corresponds to the reactant state, the
boundary b(t)can be interpreted as a time-dependent energy
barrier that needs to be surpassed to reach the product state.
Accordingly, the first-passage time can be interpreted as the
reaction time.
3 Level-crossing theory for a moving barrier
3.1 Hazard-rate representation of first-passage-time
density
To find approximations to the FPT density from approxi-
mate hazard rates, we use concepts from renewal theory,
especially the notion of hazard rate and survival probabil-
ity (Cox 1962). Because the process Eq. (5) starts at time
0, the hazard rate λ(t)is defined here as the conditional
probability per small time interval dtto find a boundary
crossing in the interval (t,t+dt)given the absence of
crossings in the interval (0,t). On the other hand, the sur-
vival probability S(t)is defined as the probability of an
absence of crossings in (0,t). The two definitions imply that
S(t+dt)=S(t)(1−λ(t)dt), hence dS(t)/dt=−λ(t)S(t).
Because the survival probability is unity at time t=0, we
thus obtain S(t)=exp −t
0λ(s)dsfor t>0. The prob-
ability to find the first crossing after time 0 in the interval
(t,t+dt)is equal to the probability to find a crossing in
(t,t+dt)and to have no crossings in (0,t). Hence, the FPT
density is given by the product P(t)=λ(t)S(t),or
P(t)=λ(t)exp −t
0
λ(s)ds.(7)
Given the hazard rate λ(t)for t>0, Eq. (7) provides a
simple formula for the FPT density. An advantage of this
representation is that the exponential factor can be turned
into a first-order differential equation,
P(t)=λ(t)S(t), dS
dt=−λ(t)S(t), S(0)=1.(8)
Thus, if the hazard rate λ(t)can be efficiently computed
for t>0, this representation permits an efficient numeri-
cal integration of the first-passage-time density forward in
time. Therefore, the main strategy in this paper is to derive
computationally efficient approximations for the hazard rate.
In general, the calculation of the hazard rate is as difficult
as the calculation of the FPT density itself. However, finding
approximations for λ(t)has several advantages over direct
approximations of P(t). Firstly, as a probability density, P(t)
must satisfy the normalization to unity. Thus, the value of the
FPT density at different times cannot be calculated indepen-
dently. In particular, the value of P(t)strongly depends on
the values for t∈(0,t). By contrast, λ(t)is not a probability
density and can thus, in principle, be arbitrary as long as it
is nonnegative and S(t)=exp −t
0λ(s)dsconverges to
zero as t→∞. Thus, if we are able to find any approxi-
mation for λ(t), the normalization of P(t)is guaranteed by
Eq. (7).
Secondly, the character of the hazard rate is more local
in time than the FPT density, and thus, we expect more
efficient approximations for the hazard rate. The non-local
character of P(t)has been already mentioned above. More-
over, the non-locality becomes particularly evident by the
integral in Eq. (7), which accumulates the history of hazard
rates. The exponential factor S(t)shaped by this integral thus
contributes a trivial history-dependence of the FPT density
P(t), which is present already for time-homogeneous pro-
cesses. By contrast, this trivial history-dependence is divided
out in the hazard rate λ(t)=P(t)/S(t).Theremaining
time-dependence of the hazard rate singles out effects of
non-stationarity and explicit time-dependence of the system,
which can be captured by local variables. Thirdly, because
of the locality in time, time-dependent rates are interesting
in its own right as they are often the natural choice to model
escape processes in terms of a Markovian dynamics and mas-
ter equations.
From the above considerations, it becomes clear that the
hazard rate representation, Eq. (7), is only useful if we suc-
ceed to derive approximations for λ(t)that are local in time.
This means that we are seeking an approximation of the haz-
ardrateintheform
λ(t)≈Φb(t), ˙
b(t),...,{zi(t)},t,(9)
which may depend on time explicitly and through a few
variables such as the value and its derivative of the time-
dependent boundary, b(t)and ˙
b(t), respectively, and possibly
through a few auxiliary variables zi(t)that obey simple ordi-
nary differential equations. Note that we use the notations Φ
for the boundary-dependent hazard rate of the model Eq. (5)
123
Loading more pages...