
Identification of the Catalytic Residues in the Cyclase
Domain of the Class IV Lanthipeptide Synthetase SgbL
Julian D. Hegemann*[a] and Roderich D. Süssmuth*[a]
Lanthipeptides belong to the family of ribosomally synthesized
and post-translationally modified peptides (RiPPs) and are
subdivided into different classes based on their processing
enzymes. The three-domain class IV lanthipeptide synthetases
(LanL enzymes) consist of N-terminal lyase, central kinase, and
C-terminal cyclase domains. While the catalytic residues of the
kinase domains (mediating ATP-dependent Ser/Thr phosphor-
ylations) and the lyase domains (carrying out subsequent
phosphoserine/phosphothreonine (pSer/pThr) eliminations to
yield dehydroalanine/dehydrobutyrine (Dha/Dhb) residues)
have been characterized previously, such studies are missing for
LanL cyclase domains. To close this gap of knowledge, this
study reports on the identification and validation of the
catalytic residues in the cyclase domain of the class IV
lanthipeptide synthetase SgbL, which facilitate the nucleophilic
attacks by Cys thiols on Dha/Dhb residues for the formation of
β-thioether crosslinks.
Representatives of the RiPP natural product family are defined
through a shared biosynthetic logic, where genetically encoded
precursor peptides are matured into the final natural products
by the activity of processing enzymes.[1]
Lanthipeptides are members of the RiPP family that contain
characteristic β-thioether crosslinks; so-called (methyl)
lanthionine ((Me)Lan) residues.[1] These crosslinks are installed
by: 1) activation/elimination of Ser/Thr hydroxy groups to yield
Dha/Dhb residues, and 2) nucleophilic attacks on these
unsaturated double bonds by Cys thiolates to yield, after an
additional protonation step, the (Me)Lan moieties.
The lanthipeptide precursors contain N-terminal leader and
C-terminal core peptide regions.[1] The leader regions feature
conserved motifs needed for substrate recognition by the
processing enzymes.[1–2] The core peptide regions are those
where the dehydroalanine/dehydrobutyrine (Dha/Dhb) residues
are intermediately formed and (sometimes only partially)
converted into the (Me)Lan crosslinks (Figure 1). After full
modification of the core peptide, the mature lanthipeptide is
released via proteolytic removal of the leader and subsequently
often exported into the extracellular space.[1–3]
Lanthipeptides can be further subclassified on the basis of
their corresponding biosynthetic enzymes and there are
currently five different classes known.[1,4] Class IV lanthipeptide
synthetases, so-called LanL enzymes, feature a three-domain
architecture consisting of a lyase, a kinase, and a cyclase
domain (Figure 1A).[1,3a,4e,5] LanLs accomplish the generation of
the Dha/Dhb residues through the concerted action of the lyase
and kinase domains. First, the central kinase domain binds the
precursor and facilitates the ATP-dependent Ser/Thr
phosphorylation.[1,2,6] Then, the N-terminal lyase domain cata-
lyzes the phosphate elimination from the pSer/pThr residues,
yielding the Dha/Dhb residues[1,7] (Figure 1B). Finally, the C-
terminal cyclase domain mediates the nucleophilic attack of the
Dha/Dhb residues by Cys thiolates and facilitates the subse-
[a] Dr. J. D. Hegemann, Prof. Dr. R. D. Süssmuth
Institute of Chemistry, Technische Universität Berlin
Strasse des 17. Juni 124, 10623, Berlin (Germany)
E-mail: [email protected]
Supporting information for this article is available on the WWW under
https://doi.org/10.1002/cbic.202100391
© 2021 The Authors. ChemBioChem published by Wiley-VCH GmbH. This is
an open access article under the terms of the Creative Commons Attribution
Non-Commercial NoDerivs License, which permits use and distribution in
any medium, provided the original work is properly cited, the use is non-
commercial and no modifications or adaptations are made.
Figure 1. (A) Schematic representation of the domain organization in class IV
lanthipeptide synthetases. (B) (Me)Lan formation by class IV lanthipeptide
synthetases. The domain active during a specific step is shown in color,
while the domains not participating in this step of catalysis are shown in
grey. (C) Schematic representation of the class IV lanthipeptide globisporin;
Abu=aminobutyric acid.
ChemBioChem
Communications
doi.org/10.1002/cbic.202100391
3169ChemBioChem 2021,22, 3169–3172 © 2021 The Authors. ChemBioChem published by Wiley-VCH GmbH
Wiley VCH Dienstag, 09.11.2021
2122 / 219836 [S. 3169/3172] 1
quent protonation of the resulting enolates to yield the (Me)Lan
crosslinks (Figure 1B).[1,2b,4e,6]
In our recent review[1b] on class III and IV lanthipeptide
synthetases, we noted that whereas the catalytic residues of the
lyase[7] and kinase[6] domains of class IV lanthipeptide synthe-
tases have been reported, the catalytic residues of the LanL
cyclase domains still remained uncharacterized. Due to the
homology between the cyclases of class I, II, and IV lanthipep-
tide synthetases, we further hypothesized that class IV cyclase
domains would feature the same conserved catalytic residues
that were previously reported[1b,c,8] for class I and II cyclases.
Indeed, an alignment of well-studied class I[1c,8a–d] and II
cyclases[1c,8b,e] with the cyclase domain of the previously
characterized class IV lanthipeptide synthetase SgbL[2b,6] as well
as with the cyclase domains of a selection of other reported
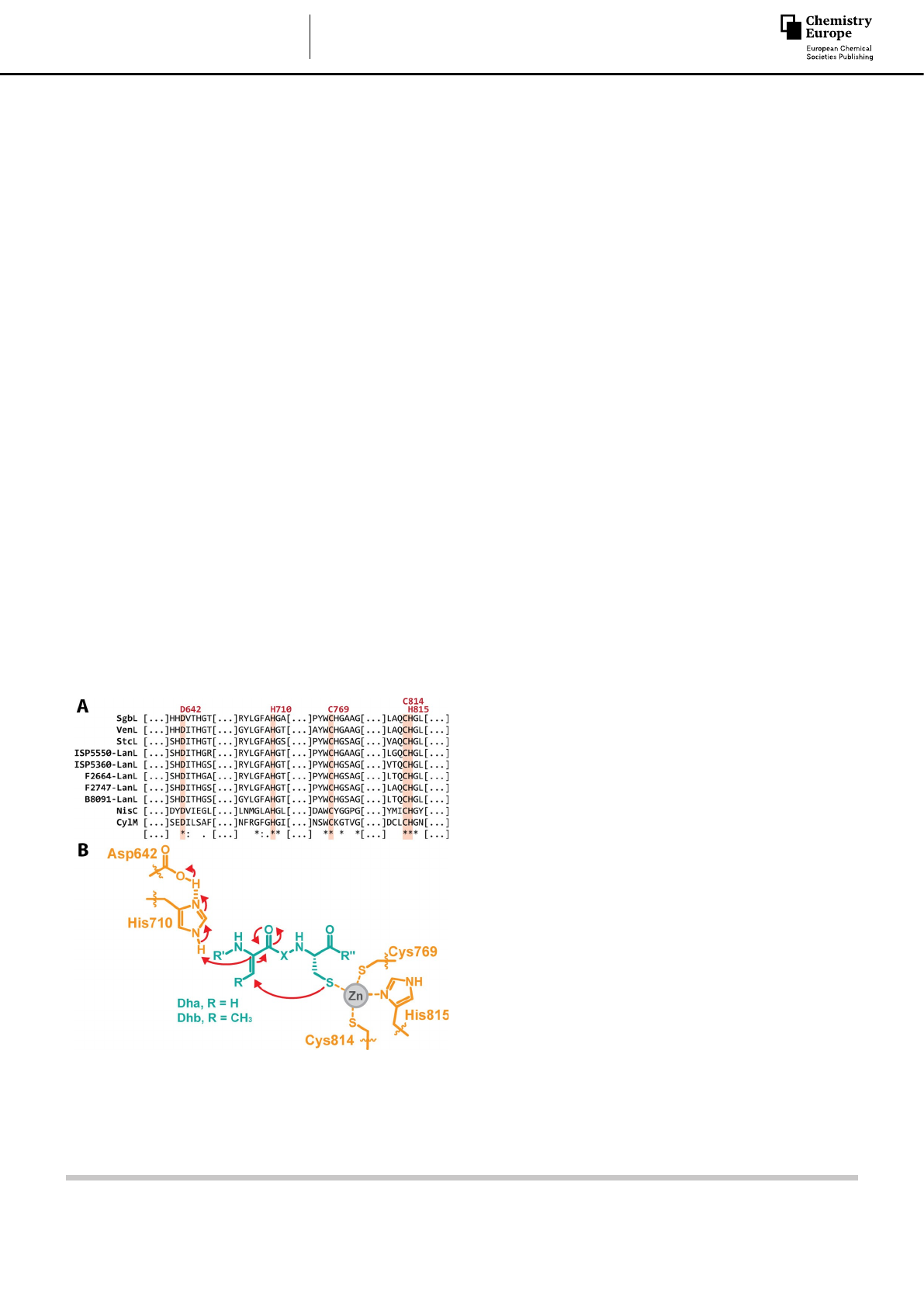
class IV enzymes[2b,3a,4e,5] (Figure 2A) shows the full conservation
of the putative catalytic residues. To experimentally validate the
role of these residues in (Me)Lan formation, we performed a
mutational analysis of the SgbL enzyme that catalyzes the core
peptide modification during the biosynthesis of the class IV
lanthipeptide globisporin (Figure 1C).[2b,6]
By mutation of the his6-sgbL gene in an expression vector,
heterologous expression in E. coli, and in vitro reconstitution,
the five predicted catalytic residues were exchanged to Ala.
According to bioinformatic analysis, three of these residues
(Cys769, Cys814, His815) would be coordinating a zinc ion that
would act as a Lewis acid to increase the nucleophilicity of the
Cys thiols in the SgbA core region (Figure 2B).[1b,c,8a–d] In addition,
His710 would fulfill the role of the catalytic acid that protonates
the enolate intermediates and that is further activated by
interaction with Asp642 (Figure 2B).[1b,c,8a–d]
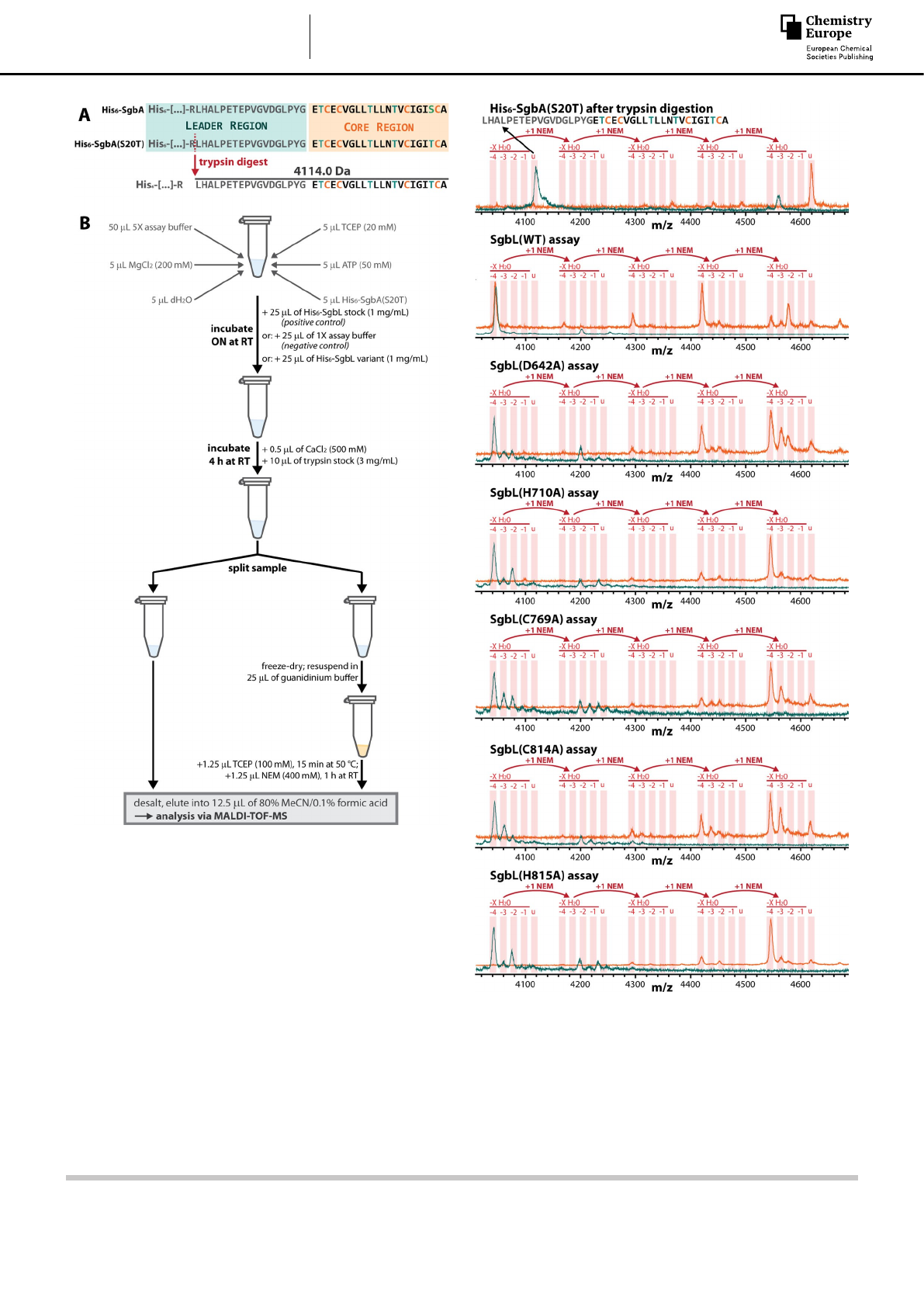
To assess if the generated SgbL variants (SgbL(D642A), SgbL
(H710A), SgbL(C769A), SgbL(C814A), SgbL(H815A)) are indeed
unable to catalyze the (Me)Lan formation, a series of in vitro
assays were performed using the SgbL WT enzyme as a positive
control. However, whereas Ser/Thr dehydrations are easily
observable by mass spectrometry (MS) due to the loss of a
water molecule (18 Da), the (Me)Lan formation is mass
neutral. Hence, the lack of (Me)Lan formation can only be
detected indirectly by labeling of the free Cys residues present.
This labeling can be accomplished by addition of N-ethyl-
maleimide (NEM), a thiol-selective electrophile.[2b,6,9] Whenever a
free thiol adds to an NEM molecule, the mass of the compound
increases by 125 Da, which can be readily tracked by MS.
Another important aspect to consider for these assays is
that the steric hindrance imposed by the methyl groups in Thr-
derived Dhb side chains efficiently suppresses non-enzymatic
cyclizations to occur, while the less sterically hindered unsatu-
rated double bonds in Dha residues can be more readily
attacked by Cys thiol nucleophiles in the absence of an
enzyme.[2b] Thus, a variant of the precursor peptide SgbA was
heterologously produced, where the only ring-forming Ser in
the core region was replaced with a Thr (His6-SgbA(S20T),
Figure 3A) to suppress the background of non-enzymatic Lan
formation in our assays.
Accordingly, the His6-SgbA(S20T) precursor variant was
incubated overnight under assay conditions either by itself
(negative control), with SgbL (positive control), or with one of
the five SbgL variants (Figure 3B).
On the next day, the reaction mixtures were digested with
trypsin to facilitate the MS analysis of the modified SgbA(S20T)
core peptides.[2b] The samples were then split to enable the
comparison of the mass spectra before and after NEM treat-
ment. Hence, one part of each sample was directly desalted and
applied to MS, while the other part was first treated with NEM
before desalting and MS analysis (Figure 3B).
Indeed, all SgbL variants introduced up to four dehydrations
into the His6-SgbA(S20T) precursor peptide as did the WT
enzyme (Figure 4), which was expected as it was
previously[2b,4e,6] shown that the LanL lyase/kinase domains can
incorporate the dehydrations independently from the cyclase
domain.
However, when comparing the results of the NEM assays, it
became apparent how the Ala exchanges affected the ability of
SgbL to introduce the MeLan crosslinks into His6-SgbA(S20T).
The assay with WT SgbL yielded a significant amount of fully-
cyclized core peptide that did not add any NEM. In contrast,
none of the SgbL variants were able to install all four MeLan
crosslinks into the precursor peptide and the major products of
all the SgbL variant assays always had four NEM molecules
added. Thus, these experiments clearly demonstrate that the
Ala exchange of any of the five catalytic residues predicted
through homology analysis (almost) completely abrogates the
ability of SgbL to introduce β-thioether crosslinks into His6-
SgbA(S20T).
Figure 2. (A) Excerpt of the alignment of a selection of known functional
LanL enzymes[2b,4e,5] with the representative cyclases of class I (NisC[8a]) and
class II (CylM[8e]) lanthipeptide synthetases. The red labels above the
alignment refer to the corresponding predicted catalytic residues in SgbL.
(B) Putative catalytic mechanism of the SgbL cyclase-mediated (Me)Lan
formation.
ChemBioChem
Communications
doi.org/10.1002/cbic.202100391
3170ChemBioChem 2021,22, 3169–3172 www.chembiochem.org © 2021 The Authors. ChemBioChem published by Wiley-VCH GmbH
Wiley VCH Dienstag, 09.11.2021
2122 / 219836 [S. 3170/3172] 1
14397633, 2021, 22, Downloaded from https://chemistry-europe.onlinelibrary.wiley.com/doi/10.1002/cbic.202100391 by Technische Universitaet Berlin, Wiley Online Library on [08/09/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
In conclusion, this study provides the first experimental
validation of the function of the predicted[1b] conserved catalytic
residues in the cyclase domain of a class IV lanthipeptide
synthetase. Adding to previous studies that were focused on
identifying and characterizing the catalytic residues situated in
the kinase[6] and lyase[7] domains of LanL enzymes, these
experiments therefore lead to a more complete understanding
of the underlying principles of class IV lanthipeptide biosyn-
thesis.
Figure 3. (A) An S20T exchange in the SgbA precursor peptide yields a core
peptide that can only form MeLan crosslinks. (B) Schematic representation of
the general workflow of the in vitro modification assays.
Figure 4. Results of the in vitro modification assays of His6-SgbA(S20T) with
SgbL and variants thereof. Traces in teal and orange show the MS data
before and after NEM labeling, respectively. For the negative control shown
on top, the precursor was incubated overnight under assay conditions in the
absence of any modification enzyme. The MS signals shown were obtained
by MALDI-TOF-MS analysis of the samples after desalting and using sinapic
acid as MALDI matrix; u=unmodified peptide.
ChemBioChem
Communications
doi.org/10.1002/cbic.202100391
3171ChemBioChem 2021,22, 3169–3172 www.chembiochem.org © 2021 The Authors. ChemBioChem published by Wiley-VCH GmbH
Wiley VCH Dienstag, 09.11.2021
2122 / 219836 [S. 3171/3172] 1
14397633, 2021, 22, Downloaded from https://chemistry-europe.onlinelibrary.wiley.com/doi/10.1002/cbic.202100391 by Technische Universitaet Berlin, Wiley Online Library on [08/09/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License

Acknowledgements
We would like to thank Dr. Chris Weise (Freie Universität Berlin)
for assistance with the MALDI-TOF-MS measurements. Open
Access funding enabled and organized by Projekt DEAL.
Conflict of Interest
The authors declare no conflict of interest.
Keywords: biocatalysis ·biosynthesis ·lanthipeptides ·natural
products ·RiPPs
[1] a) M. Montalbán-López, T. A. Scott, S. Ramesh, I. R. Rahman, A. J. van Heel,
J. H. Viel, V. Bandarian, E. Dittmann, O. Genilloud, Y. Goto, M. J. Grand-
e Burgos, C. Hill, S. Kim, J. Koehnke, J. A. Latham, A. J. Link, B. Martínez,
S. K. Nair, Y. Nicolet, S. Rebuffat, H. G. Sahl, D. Sareen, E. W. Schmidt, L.
Schmitt, K. Severinov, R. D. Süssmuth, A. W. Truman, H. Wang, J. K. Weng,
G. P. van Wezel, Q. Zhang, J. Zhong, J. Piel, D. A. Mitchell, O. P. Kuipers,
W. A. van der Donk, Nat. Prod. Rep. 2021,38, 130–239; b) J. D. Hegemann,
R. D. Süssmuth, RSC Chem. Biol. 2020,1, 110–127; c) L. M. Repka, J. R.
Chekan, S. K. Nair, W. A. van der Donk, Chem. Rev. 2017,117, 5457–5520.
[2] a) V. Wiebach, A. Mainz, R. Schnegotzki, M. J. Siegert, M. Hügelland, N.
Pliszka, R. Süssmuth, Angew. Chem. Int. Ed. 2020,59, 16777–16785;
Angew. Chem. 2020,132, 16920–16929; b) J. D. Hegemann, W. A.
van der Donk, J. Am. Chem. Soc. 2018,140, 5743–5754.
[3] a) H. Ren, C. Shi, I. R. Bothwell, W. A. van der Donk, H. Zhao, ACS Chem.
Biol. 2020,15, 1642–1649; b) S. Chen, B. Xu, E. Chen, J. Wang, J. Lu, S.
Donadio, H. Ge, H. Wang, Proc. Natl. Acad. Sci. USA 2019,116, 2533–2538.
[4] a) F. Román-Hurtado, M. Sánchez-Hidalgo, J. Martín, F. J. Ortiz-López, O.
Genilloud, Antibiotics 2021,10, 403; b) M. Xu, F. Zhang, Z. Cheng, G.
Bashiri, J. Wang, J. Hong, Y. Wang, L. Xu, X. Chen, S. X. Huang, S. Lin, Z.
Deng, M. Tao, Angew. Chem. Int. Ed. 2020,59, 18029–18035; Angew.
Chem. 2020,132, 18185–18191; c) A. M. Kloosterman, P. Cimermancic,
S. S. Elsayed, C. Du, M. Hadjithomas, M. S. Donia, M. A. Fischbach, G. P.
van Wezel, M. H. Medema, PLoS Biol. 2020,18, e3001026; d) K. Meindl, T.
Schmiederer, K. Schneider, A. Reicke, D. Butz, S. Keller, H. Guhring, L.
Vertesy, J. Wink, H. Hoffmann, M. Bronstrup, G. M. Sheldrick, R. D.
Sussmuth, Angew. Chem. Int. Ed. 2010,49, 1151–1154; Angew. Chem.
2010,122, 1169–1173; e) Y. Goto, B. Li, J. Claesen, Y. Shi, M. J. Bibb, W. A.
van der Donk, PLoS Biol. 2010,8, e1000339.
[5] D. Iftime, M. Jasyk, A. Kulik, J. F. Imhoff, E. Stegmann, W. Wohlleben, R. D.
Sussmuth, T. Weber, ChemBioChem 2015,16, 2615–2623.
[6] J. D. Hegemann, L. Shi, M. L. Gross, W. A. van der Donk, ACS Chem. Biol.
2019,14, 1583–1592.
[7] Y. Goto, A. Okesli, W. A. van der Donk, Biochemistry 2011,50, 891–898.
[8] a) B. Li, J. P. Yu, J. S. Brunzelle, G. N. Moll, W. A. van der Donk, S. K. Nair,
Science 2006,311, 1464–1467; b) X. Yang, W. A. van der Donk, ACS Chem.
Biol. 2015,10, 1234–1238; c) B. Li, W. A. van der Donk, J. Biol. Chem. 2007,
282, 21169–21175; d) M. Helfrich, K. D. Entian, T. Stein, Biochemistry 2007,
46, 3224–3233; e) S. H. Dong, W. Tang, T. Lukk, Y. Yu, S. K. Nair, W. A.
van der Donk, eLife 2015,4, e07607.
[9] a) J. D. Hegemann, S. C. Bobeica, M. C. Walker, I. R. Bothwell, W. A.
van der Donk, ACS Synth. Biol. 2019,8, 1204–1214; b) C. J. Thibodeaux, T.
Ha, W. A. van der Donk, J. Am. Chem. Soc. 2014,136, 17513–17529.
Manuscript received: August 3, 2021
Revised manuscript received: September 6, 2021
Accepted manuscript online: September 7, 2021
Version of record online: September 12, 2021
ChemBioChem
Communications
doi.org/10.1002/cbic.202100391
3172ChemBioChem 2021,22, 3169–3172 www.chembiochem.org © 2021 The Authors. ChemBioChem published by Wiley-VCH GmbH
Wiley VCH Dienstag, 09.11.2021
2122 / 219836 [S. 3172/3172] 1
14397633, 2021, 22, Downloaded from https://chemistry-europe.onlinelibrary.wiley.com/doi/10.1002/cbic.202100391 by Technische Universitaet Berlin, Wiley Online Library on [08/09/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
Loading more pages...