
molecules
Article
Immunological Analysis of Isothiocyanate-Modified
α-Lactalbumin Using High-Performance Thin
Layer Chromatography
Jenny Spöttel 1, Johannes Brockelt 1, Svenja Badekow 1and Sascha Rohn 1,2,*
Citation: Spöttel, J.; Brockelt, J.;
Badekow, S.; Rohn, S. Immunological
Analysis of Isothiocyanate-Modified
α-Lactalbumin Using
High-Performance Thin Layer
Chromatography. Molecules 2021,26,
1842. https://doi.org/10.3390/
molecules26071842
Academic Editors: Tuba Esatbeyoglu
and Banu Bayram
Received: 1 March 2021
Accepted: 22 March 2021
Published: 25 March 2021
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2021 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
1Institute of Food Chemistry, Hamburg School of Food Science, University of Hamburg, Grindelallee 117,
[email protected] (S.B.)
2Department of Food Chemistry and Analysis, Institute of Food Technology and Food Chemistry,
Technische Universität Berlin, TIB 4/3-1, Gustav-Meyer-Allee 25, 13355 Berlin, Germany
*Correspondence: r[email protected]; Tel.: +49-30-314-72583
Abstract:
Undirected modifications between food proteins and secondary plant metabolites can occur
during food processing. The results of covalent interactions can alter the functional and biological
properties of the proteins. The present work studied the extent of which covalent conjugation of
the bioactive metabolite benzyl isothiocyanate (BITC; a glucosinolate breakdown product) to the
whey protein
α
-lactalbumin affects the protein’s allergenicity. Additional to the immunological
analysis of native untreated and BITC-modified
α
-lactalbumin, the analysis of antigenic properties
of proteolytically digested protein derivatives was also performed by high performance thin layer
chromatography and immunostaining. As a result of the chemical modifications, structural changes
in the protein molecule affected the allergenic properties. In this process, epitopes are destroyed
or inactivated, but at the same time, buried epitopes can be exposed or newly formed, so that the
net effect was an increase in allergenicity, in this case. Results from the tryptic hydrolysis suggest
that BITC conjugation sterically hindered the cleavage sites for the enzyme, resulting in reduced
digestibility and allergenicity. Residual antigenicity can be still present as short peptide fragments
that provide epitopes. The desire to make food safer for allergy sufferers and to protect sensitized
individuals from an allergenic reaction makes it clear that the detection of food antigens is mandatory;
especially by considering protein interactions.
Keywords:
whey proteins; allergenicity;
α
-lactalbumin; isothiocyanates; protein modifications;
protein antigenicity; peptide antigenicity; HPTLC immunostaining; food processing
1. Introduction
Interactions between proteins and secondary plant metabolites occur frequently in
nature, and can also arise during food production and processing. The effects of covalent
protein modifications are diverse and can influence protein folding and structure as well as
various biological (hydrolysis, antigenic and antimicrobial activity), and technofunctional
and functional properties such as viscosity, gelation, foaming, solubility, emulsification,
color, odor, and taste [
1
]. From a physiological point of view, a conjugation of secondary
plant metabolites or their degradation products with dietary proteins may reduce the avail-
ability of the health-promoting secondary metabolites or decrease the bioavailability of
essential amino acids [
2
]. However, covalent addition of natural, hydrophobic, electrophilic
plant compounds is also considered a promising method to specifically affect protein func-
tionality [
3
–
6
]. In this respect, a bunch of studies have already studied the functional and
biological properties of proteins by their modification with secondary plant metabolites. For
example, it was reported that a covalent interaction of selected polyphenols with the whey
protein
β
-lactoglobulin (
β
-LG) altered its functional properties and reduced its allergenic
Molecules 2021,26, 1842. https://doi.org/10.3390/molecules26071842 https://www.mdpi.com/journal/molecules

Molecules 2021,26, 1842 2 of 19
activity [
7
]. Rade-Kukic et al. (2011) concluded that binding of isothiocyanates to
β
-LG
changed the protein’s folding and structure, improving its technofunctional properties such
as heat aggregation, foam formation, and emulsification [
3
]. Almost a handful of studies
confirmed the change in the secondary and tertiary structure of proteins as a result of
isothiocyanate (ITC) conjugation [
6
,
8
–
11
]. ITC are degradation products of glucosinolates,
which are mainly found in Brassicales plants such as broccoli, cauliflower, brussels sprouts,
and cabbage and are associated with various health-promoting properties (e.g., antibacte-
rial, anti-inflammatory, and antidiabetogenic activity) [
2
,
4
,
6
,
12
–
16
]. Due to their functional
group, isothiocyanates possess a high electrophilicity at the carbon atom, which makes a
reaction with nucleophiles conceivable. The electrophilic center of ITC reacts with thiol and
amino groups in the side chains of proteins to form thiocarbamates and thioureas [
17
–
19
].
Keppler et al. (2017) showed that covalent conjugation of allyl isothiocyanate (being a
degradation of the glucosinolate sinigrin) to a whey protein isolate significantly affected
the physicochemical properties such as charge, aggregation, surface hydrophobicity, and
secondary structural features of the protein, depending on pH value [
6
]. It was also shown
that a modification of these proteins with allyl isothiocyanate had no significant effect on
the antibacterial activity of this protein against different strains of Staphyolococcus aureus
and Eschericha coli [6].
The reason for an intensified research on whey proteins is because of their very high
nutritional quality, being due to their high content of branched, sulfur-containing, and
essential amino acids in an advantageous composition. Besides their high biological value,
whey proteins are characterized by their extraordinary functional properties and their
solubility over a wide pH value range, making them valuable food ingredients [
3
,
20
–
23
].
Whey proteins include various albumins and globulins, among which
α
-lactalbumin (
α
-
LA) is the second most common protein in cow’s milk, accounting for 2–5% [
20
].
α
-LA is a
small, acidic protein, consisting of 123 amino acids and a molecular weight of 14.2 kDa.
In addition, it is an important Ca
2+
-binding model protein and a classic example of the
molten globule state. It is a component of lactose synthase [
20
,
24
]. For example,
α
-LA and
its hydrolysates have been found to have an antihypertensive effect in hypertension [
25
], to
contribute to stress reduction [26], and to regulate cell growth [27]. It further provides an-
timicrobial [
28
] and immunostimulatory properties [
29
]. Besides to the positive properties
of α-LA, it is one of the main allergens in cow’s milk, along with β-LG and casein [30].
In general, food allergies are mostly type 1 (immediate-type) allergies that are medi-
ated by specific IgE antibodies bound on mast cells. Binding of the antibody to the antigen
activates the mast cells and stimulates them to degranulate. There is a release of histamine
and a number of other mediators of allergic inflammation [
31
]. Type I allergenicity of the
immediate reaction is a particular immunogenicity. While immunogenicity describes the
ability to induce a humoral or cellular immune response, antigenicity describes the ability
to be specifically recognized by antibodies produced as a result of an immune response to
the given substance or molecule [
32
]. A substance that is recognized by the organism as
an antigen is capable of eliciting an immune response and therefore has an immunogenic
potential, the extent of which depends, among other things, on the molecular size and
chemical structure. In the literature it is described that immunogenic substances usually
must have a molecular weight higher than 5000 Daltons and contain antigenic regions in the
secondary and tertiary structure, so-called epitopes. Generally, various proteins in food can
therefore have an allergenic potential, because the epitopes can be specifically recognized
by antibodies and subsequently trigger an allergic reaction. The extent of immunogenicity
depends on the abundance and density of the epitopes. These epitope structures of proteins
can be formed as linear (sequential and continuous) or conformational (discontinuous)
epitopes [
33
–
37
]. The latter are formed by the folding of spatial structures such as the
secondary or tertiary conformational arrangements, while sequential epitopes are short
sections of the primary structure of proteins [31,38].
Despite cow’s milk being a valuable food source for humans and especially for infants,
these food allergens can cause allergic reactions in sensitive people. However, there

Molecules 2021,26, 1842 3 of 19
is a growing desire to make food safer for allergic sufferers and to further investigate
the problem of milk allergy. As food allergens are a widespread health problem, many
investigations have been made to modify milk ingredients to reduce or eliminate their
allergenic potential [
39
–
41
]. It is well known that immune-influencing properties are
characterized by immunogenicity and antigenicity, being related to protein structures.
Therefore, it can be assumed that modifications of proteins influence the immune properties
of the native protein. In the past, the influence of various food processing such as lactic acid
fermentation, glycation, heat treatment, hydrostatic pressure and enzymatic hydrolysis
was studied [
42
]. The results showed that proteins can aggregate, denature, or bind to
lipid structures by the aforementioned technologies. They can also be glycosylated or
glycated [
43
]. Obviously, these processing-induced structural and chemical alterations of
the proteins are accompanied by a change in immunogenicity and allergenicity [
43
,
44
]. A
large part of the current state of knowledge about the influence of the food processing on
protein’s structure and function is based on numerous studies of model foods, in particular
whey proteins from cow’s milk [
45
]. Not all food processing has the potential to reduce
milk allergenicity. Increasingly, it is reported that allergenicity may be decreased, increased,
or even remain unchanged by food techniques [
46
]. Although knowledge in this field is
constantly improving, strategies to control milk allergy have not yet been satisfactorily
solved. As the processing of milk proteins can affect the protein’s structure and function
in various ways and thus, its allergenicity, this topic should be an important focus of
future research considering a change of the allergenicity of milk proteins. In addition,
there is a need for a robust and thoroughly evaluated and validated method for food
allergenicity risk assessment that considers both protein digestion and protein analysis [
47
].
In fact, many studies focused on the investigation of the covalent interaction between
whey proteins and isothiocyanates and its effect on the structural, functional, and selected
nutritional properties of the proteins [
3
,
4
,
6
,
8
–
11
], while the studies regarding the influence
on allergenic properties are insufficient [1].
However, analysis of undirected protein modifications and their implications for aller-
genicity is quite challenging. In contrast to other separation techniques, high performance
thin layer chromatography (HPTLC) offers several advantages to study the influence of
undirected posttranslational modifications that significantly affect the polarity, solubility,
and respective properties of the protein. With little effort, it is possible to use different
solvent systems, allowing a wide polarity range to be covered. This is supported by the
variety of available chromatographic stationary phases. By varying the mobile and sta-
tionary phases, HPTLC can respond quite easily and quickly to the separation problem
at hand. Biller et al. (2015) succeeded in developing a concept for the development of
solvent systems, enabling intact proteins to be analyzed by HPTLC and providing a basis
for the development of specific detections [
48
]. As the potential BITC-protein adducts
described above can heavily influence the separation behavior, HPTLC separation might
be advantageous. Even when not separated and remaining at the starting point, it is
possible to recognize the behavior of the (modified) proteins. In a high-performance liquid
chromatography (HPLC) approach, such compounds are discriminated, as they get stuck
at the beginning of the analytical column or already in the pre-column. Another advantage
of HPTLC compared to other separation techniques is the variety of compatible detection
methods. For example, a chromatogram can be evaluated with different staining reagents,
with coupling to mass spectrometry, or with a bioactivity-driven and effect-directed analy-
sis [
49
–
51
]. The combined strategy of HPTLC followed by effect-directed detection via the
specific antigen-antibody reaction, known as HPTLC immunostaining (HPTLC-IS) [
52
], can
be used to analyze the biological function of allergenicity of proteins. The direct application
of the bioassay to the analytical plate eliminates the time-consuming transfer of analytes,
such as in immunoblotting, thereby further increasing the applicability of the method.
Several studies already described antibody-based detection methods coupled to HPTLC:
Meisen et al. (2011) succeeded in detecting glycosphingolipids [
53
], while Morschheuser
et al. (2016) presented the detection of phosphorylated peptides using antibodies [
54
].
Molecules 2021,26, 1842 4 of 19
Another study by Morschheuser et al. (2017) dealt with the immunological investigation of
proteins in milk after chromatographic separation [52].
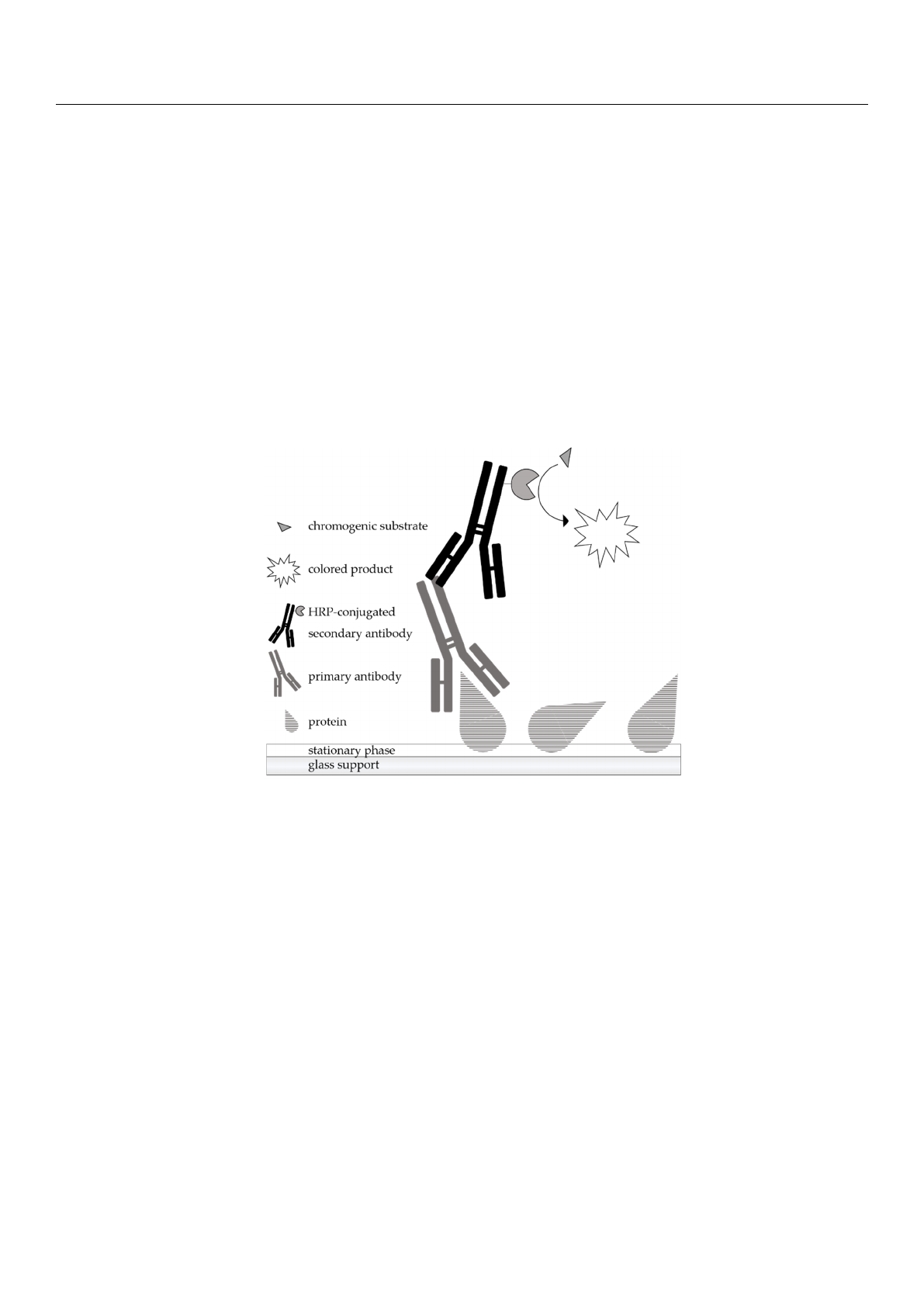
A scheme of the principle of immunostaining is shown in Figure 1. After the chromato-
graphic separation of the samples, the analytes are detected by a protein antigenicity assay
directly on the HPTLC plate. The immunological staining procedure used is very similar
to the indirect enzyme immunoassay. The post-chromatographic detection starts with a
blocking step to avoid non-specific binding of the detection antibodies on the surface of
the separation material. Afterwards, the incubation is performed with a primary antibody
that has an affinity for the target protein. The primary antibody used in the present work
was a polyclonal anti-
α
-LA antibody from the organism rabbit. The secondary antibody
needs to be specific for the host animal of the primary antibody and is conjugated with an
enzyme, here, horseradish peroxidase (HRP). For detection, an enzymatic conversion of
the chromogenic substrate tetramethylbenzidine (TMB) into a colored precipitate is finally
performed, making the first antibody visible.
Molecules 2021, 26, x FOR PEER REVIEW 4 of 19
of analytes, such as in immunoblotting, thereby further increasing the applicability of the
method. Several studies already described antibody-based detection methods coupled to
HPTLC: Meisen et al. (2011) succeeded in detecting glycosphingolipids [53], while
Morschheuser et al. (2016) presented the detection of phosphorylated peptides using an-
tibodies [54]. Another study by Morschheuser et al. (2017) dealt with the immunological
investigation of proteins in milk after chromatographic separation [52].
A scheme of the principle of immunostaining is shown in Figure 1. After the chroma-
tographic separation of the samples, the analytes are detected by a protein antigenicity
assay directly on the HPTLC plate. The immunological staining procedure used is very
similar to the indirect enzyme immunoassay. The post-chromatographic detection starts
with a blocking step to avoid non-specific binding of the detection antibodies on the sur-
face of the separation material. Afterwards, the incubation is performed with a primary
antibody that has an affinity for the target protein. The primary antibody used in the pre-
sent work was a polyclonal anti-α-LA antibody from the organism rabbit. The secondary
antibody needs to be specific for the host animal of the primary antibody and is conju-
gated with an enzyme, here, horseradish peroxidase (HRP). For detection, an enzymatic
conversion of the chromogenic substrate tetramethylbenzidine (TMB) into a colored pre-
cipitate is finally performed, making the first antibody visible.
The aim of the present work was to study adducts of the whey protein α-LA and a
bioactive metabolite from Brassicaceae vegetables, exemplarily benzyl isothiocyanate
(BITC), to subsequently estimate and compare the antigenicity of the BITC-treated and
untreated proteins. In addition to the analysis of protein derivatives, the study of residual
antigenicity of peptides after enzymatic hydrolysis, which may still carry epitopes, should
not be underestimated and was also be investigated in the present work. For this purpose,
a combined strategy of HPTLC and immunostaining was applied.
Figure 1. Schematic representation of the principle of an indirect antibody-based detection of pro-
teins after chromatographic separation (HPTLC-IS). The primary antibody having affinity for the
target protein is associated with the HRP-conjugated secondary antibody specific for the host of
the primary antibody. Visualization of the ligated antibodies is achieved by the formation of a
colored precipitate derived from a chromogenic substrate.
2. Results
2.1. HPTLC-Immunostaining (HPTLC-IS) of Intact Proteins and Its Validation
Primarily, following the work of Morschheuser et al. [52], the HPTLC method cou-
pled with antibody-based detection was applied to the whey protein α-LA and subse-
quently validated.
Figure 1.
Schematic representation of the principle of an indirect antibody-based detection of proteins
after chromatographic separation (HPTLC-IS). The primary antibody having affinity for the target
protein is associated with the HRP-conjugated secondary antibody specific for the host of the primary
antibody. Visualization of the ligated antibodies is achieved by the formation of a colored precipitate
derived from a chromogenic substrate.
The aim of the present work was to study adducts of the whey protein
α
-LA and
a bioactive metabolite from Brassicaceae vegetables, exemplarily benzyl isothiocyanate
(BITC), to subsequently estimate and compare the antigenicity of the BITC-treated and
untreated proteins. In addition to the analysis of protein derivatives, the study of residual
antigenicity of peptides after enzymatic hydrolysis, which may still carry epitopes, should
not be underestimated and was also be investigated in the present work. For this purpose,
a combined strategy of HPTLC and immunostaining was applied.
2. Results
2.1. HPTLC-Immunostaining (HPTLC-IS) of Intact Proteins and Its Validation
Primarily, following the work of Morschheuser et al. [
52
], the HPTLC method coupled
with antibody-based detection was applied to the whey protein
α
-LA and
subsequently validated.
Initially, the linear correlation of the method used was confirmed by analyzing the
standard solutions in a concentration range between 0.2
µ
g and 3
µ
g
α
-LA. The linearity
was verified by means of Mandel’s fitting test. These results obtained are comparable to

Molecules 2021,26, 1842 5 of 19
the linearity ranges of other HPTLC applications. For example, the HPTLC method of
the antimalarial substance artemisinin showed linearity in the concentration range of 0.03–
0.120
µ
g [
55
]. Reim et al. (2015) presented an HPTLC analysis of pea saponins that showed
a linear correlation in the range of 1.25–6.25
µ
g [
56
]. For proteins, Morschheuser et al. (2016,
2017) reported a linear correlation for the range of 0.1–1
µ
g lysozyme using aptamers as
detecting agents [
57
] and a linearity between 0.075–2
µ
g
β
-LG using an antibody-based
detection [52].
Additionally, in the present study, the LOD and LOQ were also determined based on
DIN standards using the calibration method (Table 1). Compared to other semi-quantitative
HPTLC analyses, the results obtained were comparable. For example, Reim et al. (2015)
calculated an LOD of 0.6
µ
g
±
17% and an LOQ of 2.1
µ
g
±
5% [
56
]. Morschheuser et al.
(2016) showed a LOD and LOQ of 0.063
µ
g
±
19% and 0.112
µ
g
±
19%, when using
HPTLC-aptastaining (HPTLC-AS) [
57
]. When using HPTLC-IS, they determined an LOD
of 0.062
µ
g
±
24% and an LOQ of 0.093
µ
g
±
22% [
52
]. The precision of the HPTLC-IS
method was within the accepted range of 15% according to the FDA guidelines.
Table 1. Statistical parameters of HPTLC-IS using anti-bovine α-LA antibodies.
Synonym Value RSD
Limit of detection 0.040 µg 0.74%
Limit of quantification 0.177 µg 3.27%
Accuracy 99.93% 4.27%
Precision 8.55% -
Coefficient of determination 0.956 1.52%
2.2. HPTLC-Immunostaining of BITC-Modified and Non-Modified α-Lactalbumin
In the following, the influence of a conjugation between
α
-LA and BITC on the al-
lergenic properties is presented. For this purpose, treated and untreated proteins were
detected after chromatographic separation using the specific antigen-antibody reaction.
Figure 2
shows a “twin plate” (one plate cut into two similar plates after the chromato-
graphic separation) stained with the protein-specific dye fluorescamine (Figure 2a), detect-
ing all proteins present. Figure 2b shows the chromatogram obtained from the immunolog-
ical staining protocol.
Molecules 2021, 26, x FOR PEER REVIEW 6 of 19
Figure 2. HPTLC analysis of BITC-α-LA derivatives as a function of the concentration of BITC. (a) Protein-specific staining
with fluorescamine (UV-light; λ = 366 nm); (b) Immunostaining with antibodies (white light) for the exclusive detection
of the antigen. (1) α-LA control (freshly prepared), (2) α-LA control (treated similar as BITC-protein derivatives), (3) BITC-
α-LA derivative “low” (cBITC = 3.8 mM), (4) BITC-α-LA derivative “moderate” (cBITC = 38 mM), (5) BITC-α-LA derivative
“high” (cBITC = 75 mM); (6) BITC-α-LA derivative “very high” (cBITC = 113 mM).
With increasing concentrations of BITC, a steady decrease in intensity of the bands
of the control (I, II and III) was observed, until they could not be detected anymore (Figure
2a,b; lanes 4-6). Similarly, a continuous increase in the intensity of new bands (green ar-
rows) was observed with increasing BITC concentration (Figure 2a,b; lanes 4–6). The new
bands had higher Rf values and appeared more intense and partially broadened. The trend
obvious from Figure 2a supported the assumption that the ITC-protein adducts formed
depending on BITC concentration to a certain extent.
In both figures, the formation of smeary streaks was pronounced at higher BITC con-
centrations (lanes 5 and 6, yellow arrows), which is probably due to an increase in the
degree of modification. It is likely that there were numerous different protein modifica-
tions formed that impaired the chromatographic separation. Moreover, after the protein-
specific staining with fluorescamine, another band can be detected attributable to an ex-
cess of residual BITC (red arrows). This identification was made by comparison with a
standard BITC solution (comparison not shown here).
2.3. HPTLC-Immunostaining of Tryptic Peptides
The present study showed that untreated and BITC-modified α-LA can be analyzed
with regard to its antigenic properties using HPTLC-IS. In addition to the native protein
structure, fragments/parts of the protein structure may also retain their antigenicity after
proteolytic hydrolysis, so that antigenic analysis of corresponding peptides must not be
neglected. Due to the low molecular weight of the peptides, their analysis by traditional
methods such as Western blotting after gel electrophoresis is limited. Consequently, an
immunological detection after HPTLC separation of the peptides resulting from the tryp-
tic digestion of α-LA was also performed in the present work. For this purpose, α-LA was
derivatized with BITC and subsequently hydrolyzed with the protease trypsin. Immuno-
logical detection by HPTLC-IS enabled the identification of the presence of residual
epitopes in the hydrolysate. Figure 3 shows the HPTLC chromatograms of tryptic pep-
tides after peptide/protein staining with fluorescamine (Figure 3a) and after im-
munostaining (Figure 3b). Antigenic and non-antigenic peptides can be differentiated.
Figure 2.
HPTLC analysis of BITC-
α
-LA derivatives as a function of the concentration of BITC. (
a
) Protein-specific staining
with fluorescamine (UV-light;
λ
= 366 nm); (
b
) Immunostaining with antibodies (white light) for the exclusive detection of
the antigen. (1)
α
-LA control (freshly prepared), (2)
α
-LA control (treated similar as BITC-protein derivatives), (3) BITC-
α
-LA
derivative “low” (c
BITC
= 3.8 mM), (4) BITC-
α
-LA derivative “moderate” (c
BITC
= 38 mM), (5) BITC-
α
-LA derivative “high”
(cBITC = 75 mM); (6) BITC-α-LA derivative “very high” (cBITC = 113 mM).
Loading more pages...