
RESEARCH ARTICLE
Perchlorate-specific proteomic stress responses of
Debaryomyces hansenii could enable microbial survival in
Martian brines
Jacob Heinz
1
| Joerg Doellinger
2
| Deborah Maus
3
| Andy Schneider
2
|
Peter Lasch
2
| Hans-Peter Grossart
4,5
| Dirk Schulze-Makuch
1,4,6,7
1
Center for Astronomy and Astrophysics, RG
Astrobiology, Technische Universität Berlin,
Berlin, Germany
2
Robert Koch-Institute, Centre for Biological
Threats and Special Pathogens, Proteomics
and Spectroscopy (ZBS6), Berlin, Germany
3
Robert Koch-Institute, Metabolism of
Microbial Pathogens (NG2), Berlin, Germany
4
Department of Plankton and Microbial
Ecology, Leibniz-Institute of Freshwater
Ecology and Inland Fisheries (IGB), Stechlin,
Germany
5
Institute for Biochemistry and Biology,
Potsdam University, Potsdam, Germany
6
GFZ German Research Center for
Geosciences, Section Geomicrobiology,
Potsdam, Germany
7
School of the Environment, Washington
State University, Pullman, Washington, USA
Correspondence
Jacob Heinz, Center for Astronomy and
Astrophysics, RG Astrobiology, Sekr. ER 3-2,
Technische Universität Berlin, Hardenbergstr.
36A, 10623 Berlin, Germany.
Email: [email protected]
Funding information
Deutsche Forschungsgemeinschaft,
Grant/Award Number: 455070607
Abstract
If life exists on Mars, it would face several challenges including the pres-
ence of perchlorates, which destabilize biomacromolecules by inducing
chaotropic stress. However, little is known about perchlorate toxicity for
microorganisms on the cellular level. Here, we present the first proteomic
investigation on the perchlorate-specific stress responses of the halotolerant
yeast Debaryomyces hansenii and compare these to generally known salt
stress adaptations. We found that the responses to NaCl and NaClO
4
-
induced stresses share many common metabolic features, for example, sig-
nalling pathways, elevated energy metabolism, or osmolyte biosynthesis.
Nevertheless, several new perchlorate-specific stress responses could be
identified, such as protein glycosylation and cell wall remodulations, pre-
sumably in order to stabilize protein structures and the cell envelope. These
stress responses would also be relevant for putative life on Mars, which—
given the environmental conditions—likely developed chaotropic defence
strategies such as stabilized confirmations of biomacromolecules or the for-
mation of cell clusters.
INTRODUCTION
Life as we know it requires energy and access to
CHNOPS (carbon, hydrogen, nitrogen, oxygen, phos-
phorus, sulfur), trace elements, and liquid water. On
Mars, energy would be provided to putative life chemi-
cally or via sunlight, carbon is accessible through the
thin but CO
2
-rich atmosphere, and other essential ele-
ments are abundant in the regolith (Clark et al., 2021).
Availability of liquid water, however, is strongly
restricted due to the low atmospheric pressure of
approximately 6 mbar and mostly subzero tempera-
tures on Mars (Martínez & Renno, 2013). One of the
few possibilities to generate liquid water in the Martian
near surface is the formation of temporarily stable
brines via deliquescence, a process in which a hygro-
scopic salt absorbs water from the atmosphere and dis-
solves within that water (Hallsworth, 2020). It has been
Received: 10 June 2022 Accepted: 27 July 2022
DOI: 10.1111/1462-2920.16152
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium,
provided the original work is properly cited.
© 2022 The Authors. Environmental Microbiology published by Society for Applied Microbiology and John Wiley & Sons Ltd.
Environ Microbiol. 2022;24:5051–5065. wileyonlinelibrary.com/journal/emi 5051

shown that deliquescent water is sufficient to drive the
metabolism of halotolerant methanogenic archaea
(Maus et al., 2020). Intriguingly, several hygroscopic
salts have been detected on Mars (Davila et al., 2010).
Among those are very deliquescent and freezing point
depressing perchlorates (ClO
4
), which are widely dis-
tributed on the Martian surface (Clark &
Kounaves, 2016) but appear in natural environments
on Earth only occasionally in hyperarid deserts (Catling
et al., 2010; Kounaves et al., 2010).
Brines formed via deliquescence provide diverse
challenges for microbial life. High salt concentrations
lead to osmotic stress and reduced water activity,
which is a measure for the amount of unbound water
molecules in a solution available for biological pro-
cesses (Hallsworth et al., 2021). Furthermore, salts can
induce ion-specific stresses like interferences with the
cell’s metabolism or changes in cell permeability
through variations in ionic hydration shells (Waajen
et al., 2020). Some anions like perchlorate additionally
evoke chaotropic stress, that is, they destabilize bioma-
cromolecules like proteins (Ball & Hallsworth, 2015),
presumably through nonlocalized attractive dispersion
forces (Hyde et al., 2017). In Pseudomonas putida,it
has been shown that chaotropic solute-induced water
stress mainly leads to upregulation of proteins involved
in stabilization of biological macromolecules and mem-
brane structure (Hallsworth et al., 2003). Furthermore,
it has been demonstrated that chaotropic effects can
be neutralized to some extent by the presence or bio-
production of kosmotropes or compatible solutes
(Bhaganna et al., 2010; Cray et al., 2015). However,
detailed research on proteomic responses to chaotropic
stress induced by perchlorate is still lacking.
Here, we present a proteomic study investigating
the perchlorate-specific stress response of Debaryo-
myces hansenii to evaluate the physiological adapta-
tions required for microorganisms to thrive in the
Martian near surface. The halotolerant yeast
D. hansenii has been chosen as a model organism as it
has been described earlier to tolerate the highest per-
chlorate concentrations reported to date (Heinz
et al., 2020,2021). This yeast provides a large meta-
bolic toolset to counteract salt stress, such as the high-
osmolarity glycerol (HOG) pathway, which enables
stress signalling and concomitant biosynthesis of glyc-
erol (Prista et al., 2016), which acts as compatible sol-
ute and antagonizes osmotic as well as chaotropic
stress (Bhaganna et al., 2016). Its close relation to the
intensively studied bakery yeast Saccharomyces cere-
visiae greatly facilitates the annotation of proteins and
thus prediction of their functions.
It has been found previously that the eukaryotic
species thus far investigated showed a higher perchlo-
rate tolerance than prokaryotes (Heinz et al., 2020).
Even though an evolutionary development of eukary-
otes on Mars might be considered unlikely due to the
relatively short habitable window of Mars, it can be
assumed that Martian microorganism—if they exist—
would have adapted over longer time scales to the
increasing aridity, salinity, and perchlorate concentra-
tions (Davila & Schulze-Makuch, 2016) and developed
defence strategies at least as efficiently and complex
as observed in D. hansenii, which is not even exposed
to high perchlorate concentrations in its natural
environment.
For the investigation of the proteome of D. hansenii,
we choose a recently developed proteomics protocol
called Sample Preparation by Easy Extraction and
Digestion (SPEED) which enables sample-type inde-
pendent deep proteome profiling with high quantitative
accuracy and precision (Doellinger, Blumenscheit,
et al., 2020; Doellinger, Schneider, et al., 2020).
This is the first study investigating perchlorate-
specific stress responses with an untargeted proteomic
approach to provide novel and fundamental under-
standing of the required cellular adaptation mecha-
nisms for life in perchlorate-rich, chaotropic habitats on
Earth, Mars, and beyond.
EXPERIMENTAL PROCEDURES
Microbial cultures
The halotolerant yeast D. hansenii (DSM 3428) was
obtained from the Leibniz Institute DSMZ—German
Collection of Microorganisms and Cell Cultures. A stock
culture was grown aerobically without shaking at 25C
(optimum growth temperature) in liquid DMSZ growth
medium #90 (3% (w/v) malt extract, 0.3% (w/v) soya
peptone) and was frequently re-inoculated. Addition-
ally, four different salt-containing liquid growth media
(DSMZ #90) were prepared having a molal (mol/kg) salt
concentration of either 1.5 mol/kg NaClO
4
, 2.4 mol/kg
NaCl, 2.4 mol/kg NaClO
4
, or 3.9 mol/kg NaCl. The latter
two concentrations (2.4 mol/kg NaClO
4
and 3.9 mol/kg
NaCl) represent the almost highest concentrations of
the respective salt enabling growth of D. hansenii. The
maximum growth-enabling concentrations reported to
date are 2.5 mol/kg NaClO
4
and 4.0 mol/kg NaCl
(Heinz et al., 2021). We choose slightly lower concen-
trations to guarantee reproducible growth of the cul-
tures and to generate sufficient biomass for protein
extraction. The other two salt concentrations (1.5 mol/
kg NaClO
4
and 2.4 mol/kg NaCl) represent moderate
salt concentrations of the respective salt (i.e. approx.
62 mol% of the respective maximum salt concentra-
tions enabling growth). The availability of two treat-
ments with the same molal salt concentrations (2.4 mol/
kg NaCl and 2.4 mol/kg NaClO
4
) allowed for an addi-
tional comparison of cellular stress responses to the
two different salt species at the same osmolality. The
growth media were prepared by mixing the media
5052 HEINZ ET AL.
components, the respective salt and water, followed by
pH adjustment (pH 5.6) and sterile filtration. All treat-
ments (no salt, 1.5 mol/kg NaClO
4
, 2.4 mol/kg NaCl,
2.4 mol/kg NaClO
4
, and 3.9 mol/kg NaCl) were inocu-
lated as biological triplicates, that is, for each treatment
three different samples were inoculated. The salt-free
treatment and the samples containing 1.5 mol/kg
NaClO
4
and 2.4 mol/kg NaCl were inoculated with the
salt-free stock culture. Hence, the two saline treatments
with moderate salt concentrations (1.5 mol/kg NaClO
4
and 2.4 mol/kg NaCl) experienced a salt shock after
inoculation. Since the respective salt shock would be
too intense in 2.4 mol/kg NaClO
4
and 3.9 mol/kg NaCl
treatments to enable growth, these samples were inoc-
ulated with long-term adapted cultures already grown
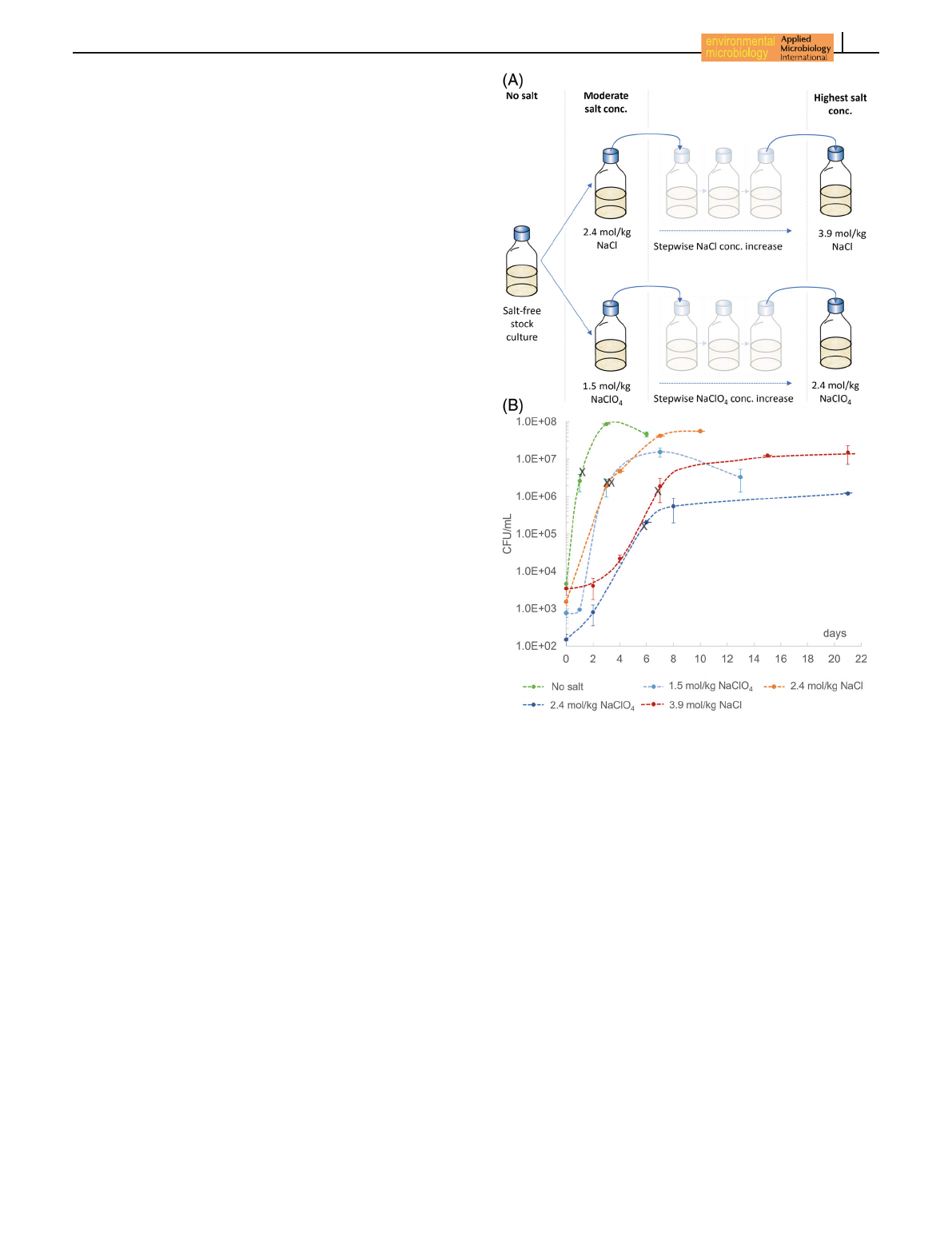
at the respective salt concentration (Figure 1A).
Sample preparation for proteomics
Protein extraction was conducted using the recently
developed filter-aided Sample Preparation by Easy
Extraction and Digestion (fa-SPEED) protocol
(Doellinger, Schneider, et al., 2020). Cells were centri-
fuged for 3 min at 5.000 gafter reaching late expo-
nential growth phase, which is approximately 1 day for
salt-free treatments, 3 days for 1.5 mol/kg NaClO
4
and
2.4 mol/kg NaCl, 6 days for 2.4 mol/kg NaClO
4
, and
7 days for 3.9 mol/kg NaCl (Figure 1B). Cell pelleting in
3.9 mol/kg NaCl samples was incomplete (turbid super-
natant) but sufficient for further protein extraction. The
reason for incomplete pelleting is presumably an
electrostatic repulsion of cells because dilution of addi-
tional test samples with water did not result in larger
pellets but gently stirring with a grounded metal rod
before centrifugation did. The cell pellets were washed
three times with phosphate buffer saline (PBS) followed
by cell lysing with 50 μl trifluoroacetic acid (TFA) for
3 min at 70C. Afterwards, samples were neutralized
with 500 μl 2 M tris(hydroxymethyl)aminomethane
(TRIS) solution. After adding 55 μl reduction/alkylation
buffer (100 mM tris(2-carboxyethyl)phosphine/400 mM
2-Chloracetamid), the samples were incubated at 95C
for 5 min.
Protein concentrations were determined by turbidity
measurements at 360 nm using GENESYS™10S UV–
Vis spectrophotometer (Thermo Fisher Scientific). The
50 μg of proteins was diluted to 40 μl using a 10:1 (v/v)
mixture of 2 M TrisBase and TFA, mixed with 160 μl
acetone and incubated for 2 min at RT. For samples
containing less than 50 μg proteins per 40 μl sample,
the volumes of sample and acetone were increased at
constant sample/acetone ratio until 50 μg protein/
sample were reached. Afterwards, proteins were cap-
tured on Ultrafree
®
-MC (0.5 ml) centrifugal devices,
0.2 μm, PTFE (Merck) at 5000 gfor 2 min. The sam-
ples were washed successively with 200 μl 80% (v/v)
acetone, 200 μl 100% acetone and 200 μl n-pentane at
5000 gfor 2 min each.
Subsequently, 40 μl of digestion buffer (50 mM
ammonium bicarbonate) containing trypsin (1:25
[enzyme-to-protein ratio] Trypsin Gold, Mass Spectrom-
etry Grade [Promega]) was added to the filter contain-
ing the proteins followed by incubation at 37C for 20 h.
The sample solution containing the digested proteins
was centrifuged at 5.000 gfor 2 min and the filter
was washed subsequently with 40 μl digestion buffer
FIGURE 1 Workflow of the inoculation procedure and
corresponding growth curves. (A) A salt-free stock culture of
D. hansenii was frequently re-inoculated into fresh growth medium.
An aliquot of this culture was used to inoculate growth media with
moderate salt concentrations (2.4 mol/kg NaCl and 1.5 mol/kg
NaClO
4
). To obtain cell growth at even higher salt concentrations, a
stepwise concentration increase was needed for each inoculation
step. The maximum salt concentrations used in this study were
3.9 mol/kg NaCl and 2.4 mol/kg NaClO
4
. (B) Growth curves of all
samples used in this study (n=3). Cells were harvested for protein
extraction in the late exponential growth phase of the respective
treatments (label with ‘X’).
PERCHLORATE-SPECIFIC MICROBIAL STRESS RESPONSES 5053

containing 0.1% (v/v) TFA. The 10% (v/v) TFA solution
was added until the pH of the samples reached approx-
imately 2. Peptides were desalted using the Pierce™
Peptide Desalting Spin Columns (Thermo Scientific)
according to the manufacture’s protocol no. 2162704.
The desalted samples were dried in a vacuum concen-
trator. The dried peptides were dissolved in 0.1% (v/v)
formic acid and quantified by measuring the absor-
bance at 280 nm using an Implen NP80 spectropho-
tometer (Implen, Munich, Germany).
Liquid chromatography and mass
spectrometry
Peptides were analysed on an EASY-nanoLC 1200
(Thermo Fisher Scientific, Bremen, Germany) coupled
online to a Q Exactive™HF mass spectrometer
(Thermo Fisher Scientific). One microgram of peptides
was separated on a PepSep column (15 cm length,
75 μm i.d., 1.9 μm C18 beads, PepSep, Denmark)
using a stepped 30 min gradient of 80% (v/v) acetoni-
trile (Solvent B) in 0.1% (v/v) formic acid (Solvent A) at
300 nl/min flow rate: 5%–11% (v/v) B in 2:49 min,
11%–29% (v/v) B in 18:04 min, 29%–33% (v/v) B in
3:03 min, 33%–39% (v/v) B in 2:04 min, 39%–95%
(v/v) B in 0:10 min, 95% (v/v) B for 2:50 min, 95%–0%
(v/v) B in 0:10 min and 0% (v/v) B for 0:50 min. Column
temperature was kept at 50C using a butterfly heater
(Phoenix S&T, Chester, PA, USA). The Q Exactive™
HF was operated in a data-independent (DIA) manner
in the m/z range of 345–1650. Full scan spectra were
recorded with a resolution of 120,000 using an auto-
matic gain control (AGC) target value of 3 10
6
with a
maximum injection time of 100 ms. The full scans were
followed by 62 DIA scans of dynamic window widths
using an overlap of 0.5 Th (Doellinger, Blumenscheit,
et al., 2020). DIA spectra were recorded at a resolution
of 30,000 using an AGC target value of 3 10
6
with a
maximum injection time of 55 ms and a first fixed mass
of 200 Th. Normalized collision energy (NCE) was set
to 27% and default charge state was set to 3. Peptides
were ionized using electrospray with a stainless-steel
emitter, I.D. 30 μm (PepSep, Denmark) at a spray volt-
age of 2.1 kV and a heated capillary temperature
of 275C.
Data analysis and statistical information
Protein sequences of Debaryomyces hansenii
(UP000000599, downloaded 16/10/20), were obtained
from UniProt (UniProt Consortium, 2019). A spectral
library was predicted for all possible peptides with strict
trypsin specificity (KR not P) in the m/z range of 350–
1150 with charge states of 2–4 and allowing up to one
missed cleavage site using Prosit (Gessulat
et al., 2019). Input files for library prediction were gen-
erated using EncyclopeDIA (Version 0.9.5) (Searle
et al., 2018). The data were analysed using the pre-
dicted library with fixed mass tolerances of 10 ppm for
MS
1
and 20 ppm for MS
2
spectra using the ‘robust LC
(high accuracy)’quantification strategy. The false dis-
covery rate was set to 0.01 for precursor identifications
and proteins were grouped according to their respec-
tive genes. The resulting pg_matrix.tsv file was used
for further analysis in Perseus (Version 1.6.5.0)
(Tyanova et al., 2016).
The same programme was used to z-normalize pro-
tein abundances followed by ANOVA (FDR =0.01)
and post hoc testing (FDR =0.05). Subsequently, the
abundances of biological triplicates were median aver-
aged, and the relative log2-fold changes of the salt-
containing (saline) treatments compared to the salt-free
control were calculated. The results were filtered for
significant pairs of the salt-free samples and at least
one of the saline treatments and were then plotted into
a hierarchical clustered heatmap. Additionally, volcano
plots have been generated with the same software after
t-test of the z-normalized protein abundances. Protein
groups of interest were annotated and analysed with
the STRING database (https://string-db.org/)
(Szklarczyk et al., 2021) regarding enriched metabolic
pathways and the formation of functional protein
clusters.
RESULTS
In order to distinguish the perchlorate-specific stress
response of D. hansenii to the stress caused by NaCl,
proteomes of cell cultures containing either NaClO
4
,
NaCl or no additional salts in growth medium DSMZ
#90 were analysed. Two different salt concentration
regimes were investigated (Figure 1A). At moderate
salt concentrations (1.5 mol/kg NaClO
4
and 2.4 mol/kg
NaCl), growth was obtained by inoculation with a salt-
free culture to provoke a salt shock response. How-
ever, the highest salt concentrations used in this study
(2.4 mol/kg NaClO
4
and 3.9 mol/kg NaCl) only enabled
growth when cells were long-term adapted to stepwise
increasing salt concentrations (Figure 1A). All samples
were prepared as biological triplicates and cells were
harvested in the late exponential growth phase in order
to obtain sufficient biomass for protein extraction
(Figure 1B). It should be noted that while 2.4 mol/kg
NaClO
4
is already close to the growth-limiting NaClO
4
concentration (similar growth rate to 3.9 mol/kg NaCl),
2.4 mol/kg NaCl represents a readily feasible NaCl con-
centration with a growth rate similar to 1.5 mol/kg
NaClO
4
.
In total, 2713 proteins were detected representing a
bulk coding sequence coverage of approximately 43%.
Through analysis of variance (ANOVA, FDR ≤0.01) of
5054 HEINZ ET AL.
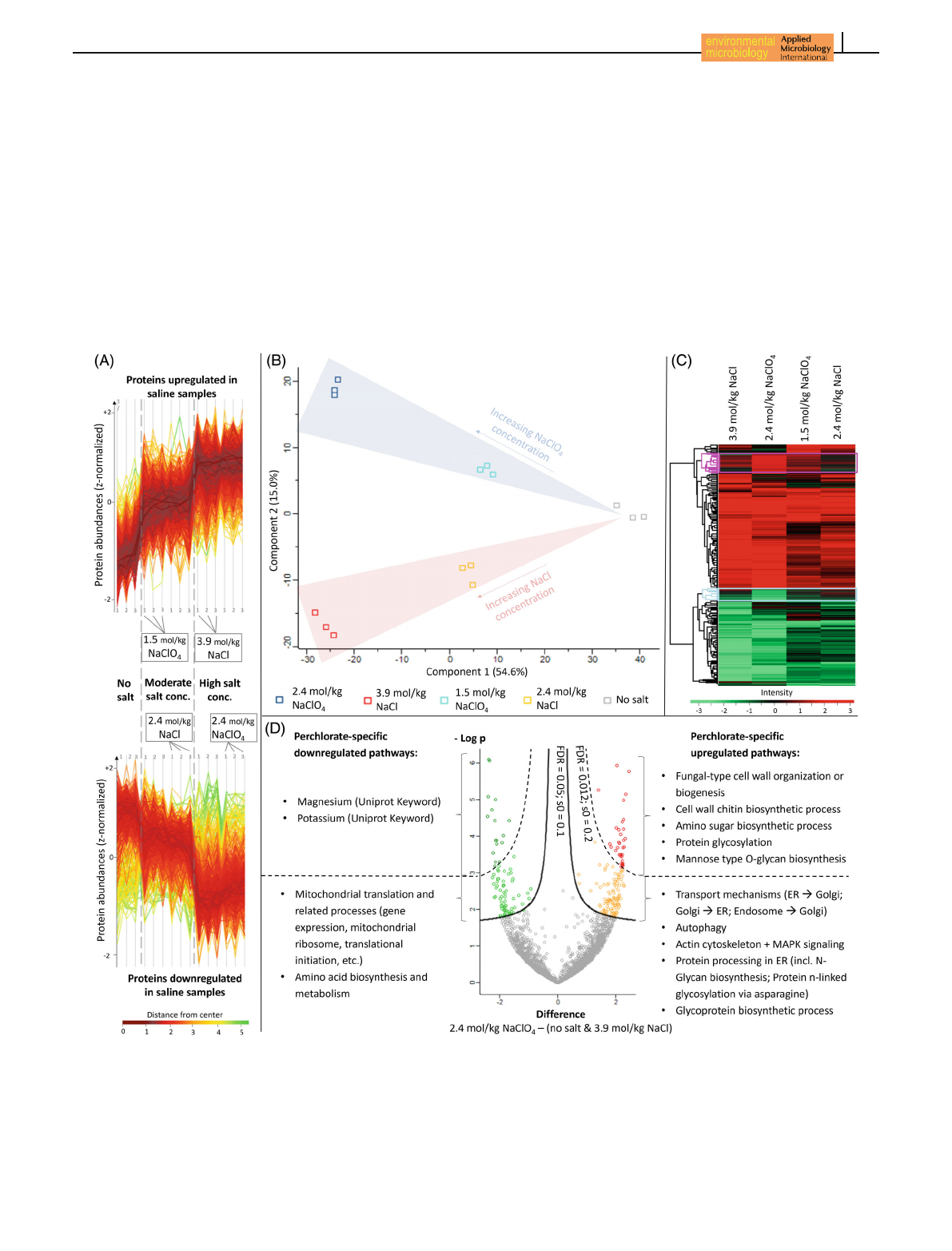
the z-normalized protein abundances, the expression
of 1099 proteins was found to be significantly different
between the five different treatment types (one salt-free
control and four salt-exposed treatments). The salt con-
centration (moderate vs. high) had a stronger impact on
the intensity of protein expression than the type of
anion as can be seen from the comparison of protein
abundances of all replicates, which show similar protein
expressions for the same salt concentration regimes
(Figure 2A). This is confirmed by the principal compo-
nent analysis (PCA), which revealed a clear clustering
of the replicates of each treatment in dependence on
salt concentration and type of anions (Figure 2B). While
the physiological response to different salt concentra-
tions clustered along principal component 1 and
explains 55% of the observed differences, the salt spe-
cies had a lower impact on the variability (15%), as
treatments exposed to chloride or perchlorate spread
along the principal component 2.
Post hoc testing (FDR ≤0.05) revealed 1068 pro-
teins to be significantly regulated in at least one of the
salt-exposed samples compared to the salt-free treat-
ment. The log2-fold changes of these proteins in the
saline treatments compared to the salt-free control
were plotted in a heatmap with upregulated proteins
coloured red and downregulated protein shown in
FIGURE 2 Results of the proteomic analyses. (A) Abundances of upregulated (upper plot) and downregulated (lower plot) proteins
expressed in all investigated samples (three replicates for each treatment as indicated in the space between the two plots). (B) Principal
component analysis (PCA) demonstrating clear clustering of all biological triplicates in dependence of salt concentration and type of anion.
(C) Heat map including all proteins passing ANOVA (FDR ≤0.01) and post hoc test (FDR ≤0.05) generated by the Perseus software after
hierarchical clustering. Upregulated proteins (compared to the salt-free treatment) are coloured red and downregulated proteins are shown in
green. Two exemplarily perchlorate-specific clusters are highlighted in pink for upregulated and in cyan for downregulated proteins. (D) Volcano
plot visualizing perchlorate-specific regulated proteins with a high (FDR ≤0.012) and a lower significance (0.012 ≤FDR ≤0.05). Significantly
regulated metabolic pathways were analysed with the STRING database.
PERCHLORATE-SPECIFIC MICROBIAL STRESS RESPONSES 5055
Loading more pages...