
This journal is © The Royal Society of Chemistry 2022 Soft Matter, 2022, 18, 6179–6191 | 6179
Cite this: Soft Matter, 2022,
18, 6179
Emergent collective dynamics of pusher and
puller squirmer rods: swarming, clustering,
and turbulence†
Arne W. Zantop * and Holger Stark *
We study the interplay of steric and hydrodynamic interactions in suspensions of elongated micro-
swimmers by simulating the full hydrodynamics of squirmer rods in the quasi two-dimensional geometry
of a Hele-Shaw cell. To create pusher or puller-type squirmer rods, we concentrate the surface
slip-velocity field more to the back or to the front of the rod and thereby are able to tune the rod’s
force-dipole strength. We study a wide range of aspect ratios and area fractions and provide
corresponding state diagrams. The flow field of pusher-type squirmer rods destabilizes ordered
structures and favors the disordered state at small area fractions and aspect ratios. Only when steric
interactions become relevant, we observe a turbulent and dynamic cluster state, while for large aspect
ratios a single swarm and jammed cluster occurs. The power spectrum of the turbulent state shows two
distinct energy cascades at small and large wave numbers with power-law scaling and non-universal
exponents. Pullers show a strong tendency to form swarms instead of the disordered state found for
neutral and pusher rods. At large area fractions a dynamic cluster is observed and at larger aspect ratio a
single swarm or jammed cluster occurs.
1 Introduction
Microscopic unicellular organisms make up a major fraction of
all life forms on our planet. They are involved in important
natural processes such as photosynthesis
1–3
or industrial pro-
cesses such as the production of enzymes
4,5
or biofuels,
6–8
or
the recycling of wastewater.
9,10
Especially in photosynthesis,
commonly viewed as the basis of life, unicellular phytoplankton
accomplish about half of the worldwide natural turnover.
2,3
However, still many aspects of the behavior of unicellular
organisms is not completely understood. In fact, many of these
life forms are self-propelling microswimmers,
11–13
as, for exam-
ple, the algae C. reinhardtii
14,15
or the bacterium E. coli.
16–18
From the perspective of physicists, their active motion gives rise
to very interesting new collective phenomena. In this article we
study the rich emergent collective dynamics of rod-shaped
model microswimmers, which we can tune between the pusher
and puller type.
Active motion is always performed in non-equilibrium and,
therefore, gives rise to new and interesting phenomena. For
example, specially designed boundaries can rectify the random
motion of active particles,
19–23
or under gravity active particles
develop polar order and even show inverted sedimentation
profiles when they are bottom-heavy.
24,25
In addition, micro-
swimmer suspensions are subject to long-range hydrodynamic
interactions with characteristic power-law decay.
13,26,27
But also
short-range steric interactions play an important role, in parti-
cular, for elongated particles, which align along each other.
12
The combination of these interactions gives rise to numerous
interesting dynamic patterns. Common examples are the for-
mation of swarms or flocks,
28–32
convection rolls and plumes,
25
fluid pumps,
33,34
vortices,
35–38
active nematic patterns,
39–41
and
the emergence of the so-called active turbulence,
42,43
termed in
analogy to classical inertial turbulence.
44,45
However, contrary to
classical turbulence, where fluid flow is driven on the macroscopic
scale, active turbulence is generated at the microscopic scale of
the self-propelled particles and then energy is dissipated on larger
scales. This mechanism causes a characteristic length scale for
the formation of vortices and patterns, which is in contrast to
the scale invariance of classical turbulence.
46–48
In particular, the
specific model parameters now determine the scaling of the
velocity power spectrum, which is no longer universal.
48–50
Active
turbulence is found for microswimmers,
51,52
active bio-filaments,
which exist in the cytoplasm,
39,53
and in growing tissue.
54,55
Institut fu
¨r Theoretische Physik, Technische Universita
¨t Berlin, Hardenbergstraße 36,
†Electronic supplementary information (ESI) available: We provide eight videos
of different collective dynamic states in the top view. Videos 1–3 show pusher rods
in the turbulent (1), single swarm (2) and dynamic cluster state (3). Videos 4–8
show puller rods in the swarming (4 and 8), single swarm (5), jammed cluster (6),
and dynamic cluster state (7). See DOI: https://doi.org/10.1039/d2sm00449f
Received 8th April 2022,
Accepted 30th June 2022
DOI: 10.1039/d2sm00449f
rsc.li/soft-matter-journal
Soft Matter
PAPER
Open Access Article. Published on 05 July 2022. Downloaded on 10/27/2022 10:58:01 AM.
This article is licensed under a
Creative Commons Attribution 3.0 Unported Licence.
View Article Online
View Journal
| View Issue
6180 | Soft Matter, 2022, 18, 6179–6191 This journal is © The Royal Society of Chemistry 2022
On the theoretical side, the dynamics of active particles has
been investigated at different levels of description. While the
celebrated Vicsek model
56–58
uses coarse-grained alignment
rules, also models with explicit steric interactions exist.
42,51,59–64
Thesemodelshavealsobeengeneralizedtoactivefilaments
31,65,66
and models, which implicitly include hydrodynamic inter-
actions.
67,68
Models, which directly simulate fluid flow and
thereby explicitly include hydrodynamic interactions, use the
method of multi-particle collision dynamics (MPCD)
25,30,32,69–79
or the lattice-Boltzmann method.
80–83
Continuum models
combine elements of the Toner-Tu
84
and Swift-Hohenberg
85
equations to generate hydrodynamic equations for active
suspensions.
46,86,87
Microswimmers are also distinguished by
their swimming mechanisms and the flow field, they generate
in the surrounding fluid.
11,13
While some microorganisms
propel with cilia located all over their surface, common bacteria
and algae propel with flagella that extent from the front or back
of the cell body. In the first case, the flow field of a source
dipole is realized, which decays as r
3
, while bacteria and algae
are termed pusher or puller-type swimmers that generate a
long-range force-dipole flow field, which decays as r
2
.
13,27
The
specific form of these hydrodynamic multipole flow fields deter-
mines the collective dynamics of the microswimmers.
32,69,74,88–90
In this context, we proposed in ref. 76 the squirmer rod as
a realistic microscopic model for elongated microswimmers.
It consists of overlapping squirmers and thereby extends the
well-known spherical squirmer model for ciliary propulsion
26,91
and its implementation in MPCD.
92
Restricting the surface slip-
velocity to the front or the back of the squirmer rod, puller and
pusher-type squirmer rods can be realized, respectively. The
hydrodynamic flow field is simulated using an efficient imple-
mentation of MPCD with a reduced compressibility such that
also collective dynamics in large and dense systems can be
studied.
93
Most recently, we presented the state diagram of
neutral squirmer rods and identified with increasing area
fraction and depending on the aspect ratio of the rods the
disordered state, dynamic swarms, a single swarm, and ultimately
ajammedcluster.
94
In this article we generalize the squirmer-rod model to
pushers and pullers with tunable force-dipole strength in the
flow field. Compared to spheroidal squirmers it has the advan-
tage that it better approximates real rodlike microswimmers
such as E. coli. Using this model, we provide a comprehensive
study of the state diagram for hydrodynamically interacting
microswimmers over a wide range of aspect ratios, densities,
and force-dipole strengths as the state diagrams in Fig. 3(a),
7(a) and 9(a) show. Hereby, we go well beyond previous works,
which focused mainly on more dilute systems
95,96
or a single
aspect ratio.
97
Thus, our work provides an overall view how
hydrodynamics and shape determine the dynamic states of
microswimmers. For example, our particle-based model nicely
illustrates that the active turbulent state occurs as a compro-
mise between the disordering hydrodynamic pusher–pusher
interactions and aligning steric interactions. We also demon-
strate that the two distinct energy cascades at low and large
wave numbers in the power spectral density of velocity
fluctuations exhibits non-universal exponents. Moreover, we
find a dynamic cluster state at large densities. Besides this
state all other states are also found in dry active rods.
98
But, in
our case, they have a specific contribution from hydrodynamics
as, for example, the turbulent and swarming states show. The
overall appearance of the states in our state diagrams can be
summarized as follows. For pushers we observe that the swarm-
ing states of neutral squirmer rods are destabilized. Instead, for
smaller aspect ratio between the disordered and dynamic
cluster state, we observe the turbulent state as already men-
tioned. At high aspect ratios, where steric interactions become
more relevant, we recover the single swarm and jammed cluster
state of neutral squirmer rods. For pullers, hydrodynamic
interactions stabilize the swarming state even for our smallest
area fraction. Thus, compared to neutral squirmer rods, states
are shifted towards lower densities. Variation of the force-dipole
strength at constant aspect ratio supports all these findings.
The article is organized as follows. In Section 2 we give a
brief overview of the methods used in this paper. Section 3
provides a detailed study of the different dynamic states of the
squirmer rods as a function of their area fraction, aspect ratio,
and force-dipole strength. We end with a summary and con-
clusions in Section 4.
2 System and methods
We first introduce the squirmer-rod model and then summarize
some details of the method of multi-particle collision dynamics,
which we use to simulate the flow fields generated by the
squirmer rods.
2.1 Model of the squirmer rod
To model shape-anisotropic microswimmers, we employ the
squirmer rod model as introduced in our previous work.
76
Squirmer rods consist of N
sq
overlapping spherical squirmers
of radius R
sq
, arranged on a line to form a single rigid body [see
Fig. 1(a)]. By varying the distance dbetween neighboring
squirmers, we can tune the aspect ratio of the squirmer rod,
a=l
S
/2R, where l
S
is the rod length. However, we do not exceed
a distance of dE0.8Rso that the surface of the rod is
sufficiently smooth. With a number of N
sq
= 10 squirmers in
this work, this amounts to a maximum aspect ratio of aE5,
which closely resembles the aspect ratio of bacteria such as
E. coli or B. subtilis.
The squirmer rods propel through the axisymmetric and
tangential slip velocity field at the surface of individual sphe-
rical squirmers,
v
s
=B
s
1
[(e
ˆxˆ
s
)xˆ
s
e
ˆ], (1)
which is imposed on the surrounding fluid.
26,92
Here, e
ˆis the
rod axis and xˆ
s
the unit vector pointing from the center of a
squirmer to a point on the squirmer surface. This generates a
source-dipole flow field, which is a higher-order singular
solution of the Stokes equations Zr
2
v=rptogether with the
incompressibility condition rv= 0, that govern fluid flow at
Paper Soft Matter
Open Access Article. Published on 05 July 2022. Downloaded on 10/27/2022 10:58:01 AM.
This article is licensed under a
Creative Commons Attribution 3.0 Unported Licence.
View Article Online
This journal is © The Royal Society of Chemistry 2022 Soft Matter, 2022, 18, 6179–6191 | 6181
the microscale. Here, vand pare the respective fluid velocity
and pressure fields, and Zis the dynamic shear viscosity. The
strength B
s
1
controls the swimming velocity v
0
= 2/3B
s
1
of the
spherical squirmer and hence of the squirmer rod. In ref. 76
we showed that the swimming velocity of the squirmer rods
vE1.2v
0
slightly exceeds the velocity of a single spherical
squirmer. Additionally, the velocity of the rods varies by 10% in
the range of aspect ratios used in this article. Although the
increased velocity at the larger aspect ratios might augment
clustering, we assume the effect to be negligible.
As described so far, the surface slip velocity of the squirmer
rod resembles ciliated microorganisms such as Paramecium.
In this realization, the profile of the slip velocity generates a
flow field in the surrounding fluid, the far field of which can be
described by a source dipole and an additional force quadru-
pole singularity, which both decay with |u
sd
|, |u
fq
|Br
3
,aswe
show in ref. 76.
However, other prominent microswimmers such as E. coli
bacteria or Chlamydomonas algae propel by rotating or beating
flagella that extent from the back or the front of their bodies,
respectively. These modes of propulsion create a pair of oppos-
ing forces that generate the more long-ranged force-dipole flow
field |u
fd
|Br
2
. To generalise the squirmer rod model to these
pusher and puller-type microswimmers, we concentrate the
surface slip velocity either to the back or the front of the
squirmer rod [see Fig. 1(a)]. This is done by multiplying
the surface flow field with the envelope function
fðx
s^
e;wÞ¼1þwtanhð10x
s^
e=lSÞ;(2)
where x
spoints from the rod center to a location on the rod
surface. The parameter wA[1, 1] determines the swimmer
type and force-dipole strength, such that for wo0 a pusher-
type swimmer is realised and likewise a puller-type swimmer
for w40. For either w=1orw= 1 this modification leads to a
completely passive half of the rods [see Fig. 1(a)], while for
intermediate values the relative contributions of the source
dipole and force dipole to the flow field vary. For w= 0, the
model again resembles the neutral squirmer rod.
In the present work, we consider the collective dynamics of
squirmer rods confined between two parallel walls. In this
so-called Hele-Shaw cell the radial decays of hydrodynamic
multipoles are modified compared to the bulk fluid, such that
theconfinedsourcedipoleandforcedipoledecayas|u
˜
sd
|Br
2
and |u
˜
fd
|Br
3
, respectively, where ris the polar distance
76,99
(see
appendix of ref. 99). As a consequence, the source dipole has the
longest range in the flow field and ultimately dominates the far
field. However, already in our previous work we realized that a
distance of Dz=6Rbetween the walls, which we will use in our
simulations, alters the relative strength of the source and force
dipoles.
76
As a consequence, the force dipole dominates the flow
field at short and medium distance as we will demonstrate in
Section 3.2.
2.2 Method of multi-particle collision dynamics
To model the fluid flow in our simulations, we employ the
meso-scale simulation technique of multi-particle collision
dynamics (MPCD).
100–102
The MPCD method is particularly
suited for solving the Navier–Stokes equations at the micro-
scale, because it includes thermal fluctuations and is straight-
forward to implement boundary conditions for complex
geometries.
The MPCD method uses a sequence of streaming and
collision steps of the point-like fluid particles. In the streaming
step the fluid particles move ballistically with their velocities
during time step Dt. Then, the simulation box is divided into
cubic cells with edge length a
0
and the velocities of the fluid
particles in one cell are modified randomly but keeping the
mean velocity or linear momentum fixed. In this work, we use a
collision rule optimized to achieve a low compressibility of the
fluid.
93
As in other MPCD methods, it includes angular
momentum conservation and a thermostat. By choosing the
MPCD fluid density n
0
= 20/a
03
,i.e., on average 20 fluid particles
per cell, and the time step Dt¼0:005a0ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
m0=kBT0
p, we obtain a
fluid viscosity of Z¼16:05 ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
m0kBT0
p=a0
2, which is comparable
to previous work.
74,76,96
Here, m
0
is the mass of one fluid
particle.
The immersed squirmer rods are modeled with the mass
density of the fluid r
0
=m
0
n
0
. The radius of the constituting
squirmersischosenasR
sq
=3a
0
and the strength of the surface slip
velocity as B
s
1
=0.1,whichresultsinaPe
´clet number of Pe E350.
94
Steric repulsion is realized with a Weeks–Chandler–Andersen
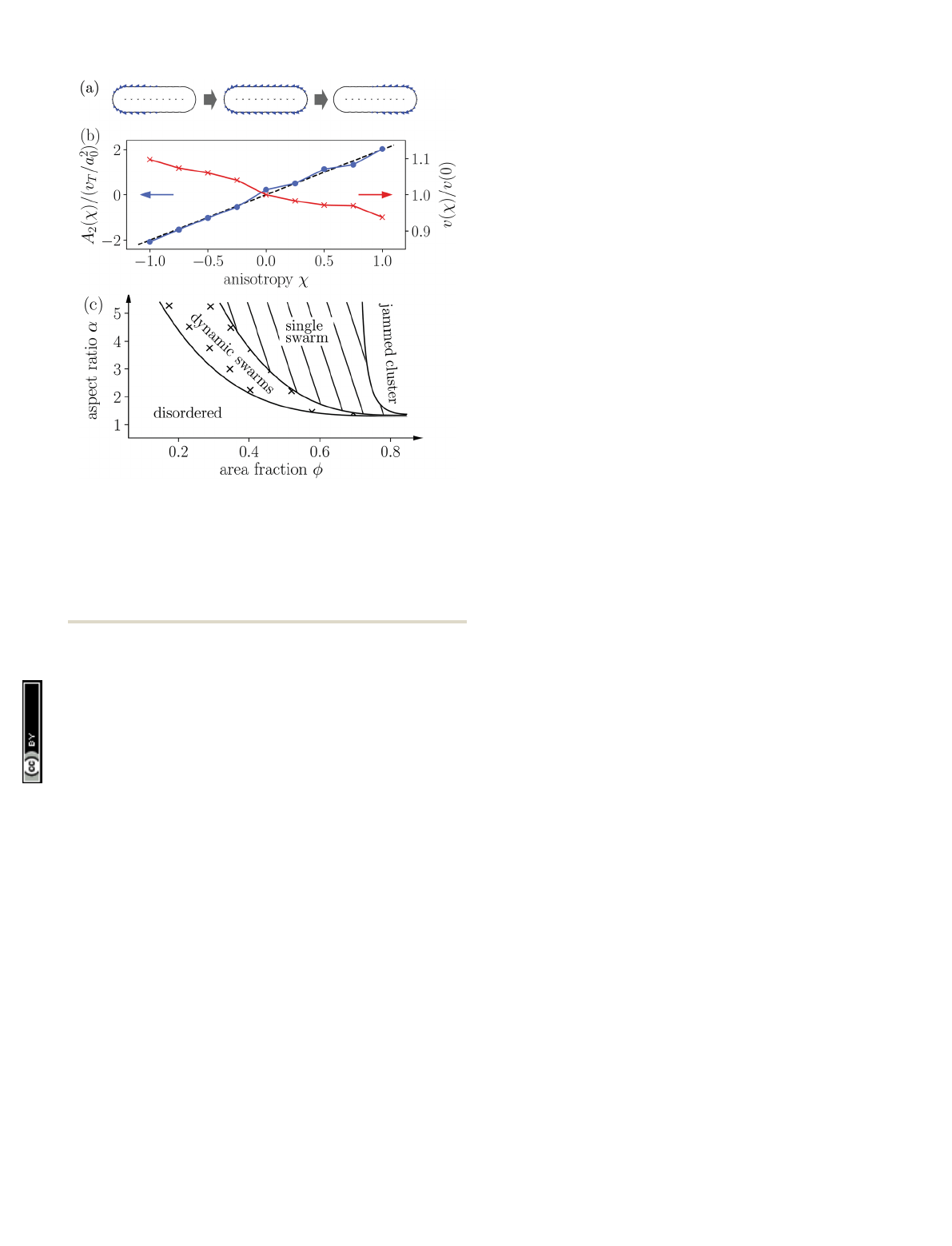
Fig. 1 (a) Sketch of the surface slip velocity of three squirmer rods with
respective swimmer-type parameters w=1, 0, and 1 (from left to right).
The arrows within the squirmer rods indicate the swimming direction e
ˆ.
The parameter wA{1, 1} can be used to smoothly vary the type of the
squirmer rod from pusher to neutral to puller. (b) Force-dipole coefficient
A
2
(w) (blue) and active velocity v(w)/v(0) (red) as a function of the swimmer
parameter w. (c) Schematic state diagram for neutral squirmer rods adapted
from ref. 94.
Soft Matter Paper
Open Access Article. Published on 05 July 2022. Downloaded on 10/27/2022 10:58:01 AM.
This article is licensed under a
Creative Commons Attribution 3.0 Unported Licence.
View Article Online
6182 | Soft Matter, 2022, 18, 6179–6191 This journal is © The Royal Society of Chemistry 2022
potential
103
that acts between two squirmers of different rods.
To ensure that there is no significant overlap between two
squirmer rods, we choose a strong force constant e
WCA
E10
4
k
B
T.
The squirmer rods move because they acquire momentum
from the surrounding fluid. In the MPCD streaming step, linear
and angular momentum is transferred to the squirmer rod by
collisions with the fluid particles. We achieve this by applying
the so-called bounce back rule, which we also modify accord-
ingly to implement the slip velocity in eqn (1) on the surface of
the squirmer rods.
100,101
Squirmer rods also contain so-called
‘‘ghost’’ particles, which improve the implementation of no-slip
boundary conditions.
101
During the MPCD collision step, they
exchange momentum with the fluid, which is ascribed to the
squirmer rod. Lastly, the equations of motion for the squirmer
rods are integrated with a refined time step of dt
MD
=Dt/3
using a symplectic splitting algorithm for rigid body molecular
dynamics.
104
For our simulations we use two different geometries.
To determine how the force dipole strength A
2
depends on
the swimmer-type parameter w, we use a cubic box of linear size
L= 100a
0
with periodic boundary conditions along all spatial
directions. In this case we use the time 10
4
Dtto equilibrate the
MPCD fluid flow fields and then average the fluctuating flow
fields over additional 5 10
5
Dttime steps during simulations.
To simulate the collective dynamics of the squirmer rods, we
consider the quasi two-dimensional geometry of a Hele-Shaw
cell of linear size L= 300a
0
in the xand ydirection. Along the
zdirection the system is confined by walls separated by a
distance Dz=6Rto mimic the experimental setups using
microfluidic chambers
51,105
or liquid–oil interfaces.
106,107
It has
also been used in previous work.
94,97
Additionally, this realization
guarantees a strong contribution of the force-dipole interaction
in the near field, as we will show in Section 3.2 and Fig. 2. In the
xand ydirection periodic boundary conditions are employed.
In this geometry, the Nsquirmer rods are confined to only
move in the midplane of the Hele-Shaw cell by a strong
harmonic potential. The rods’ initial positions are generated
randomly for area fractions f=NA
sw
/L
2
o0.6, where A
sw
is the
two-dimensional cross section of one squirmer rod. For fZ0.6
squirmer rods are placed on a rectangular lattice, all with a
randomly chosen orientation either parallel or anti-parallel to
one of the major axis of the unit cell. We simulate for a time of
10
7
Dtwhile saving snapshots every 2500Dtfor further analysis.
To ensure that the system is equilibrated, we omit the first
100 snapshots from the analysis.
To improve statistics in the study of the emergent turbulent
patterns of pusher-type squirmer rods, we perform two addi-
tional simulation runs for all turbulent states and their neigh-
boring points in the (a,f) parameter space. Furthermore, for all
these cases we also perform three simulation runs with an
increased system size of L= 600a
0
to investigate finite-size effects.
3 Results
In the following we report on our simulation results. First, we
show that the anisotropy parameter wof the surface slip-velocity
field is directly proportional to the strength of the hydro-
dynamic force-dipole field and we illustrate the flow field of a
single pusher-type squirmer rod in the Hele-Shaw cell. Then, we
thoroughly discuss the state diagrams of the strongest pusher
rod (w=1) and the strongest puller rod (w= 1) depending on
aspect ratio aand area fraction f. We describe the different
states using the velocity pair-correlation function, the power
spectral density of the velocity fluctuations, and the orienta-
tional autocorrelation function. Finally, for a specific aspect
ratio a, we show the state diagram in the space of wversus f.
3.1 Variation of the swimmer-type parameter v
To extract the force-dipole coefficient A
2
from the flow field of
the squirmer rod in the 3D bulk fluid, we follow the method
described in detail in our previous article ref. 76. To do so, we
consider the expansion of an axisymmetric flow field into a
series of hydrodynamic multipoles u(r)=u
FD
(r)+u
SD
(r)+
u
FQ
(r)+..., where the leading-order multipoles are the force
dipole, source dipole, and force quadrupole, respectively. The
radial velocity component with the general form urðr;yÞ¼
P
1
n¼1
AnrnþBnrn2
Pnðcos yÞis measured from the simula-
tions and then projected on the second Legendre polynomial
P
2
(cosy). From the resulting polynomial 5
2Ðp
0urðr;yÞP2ðcos yÞ
sin ydy¼A2r2þB2r4, we determine the force-dipole
coefficient A
2
by a polynomial fit in r
1
.
Indeed, we find a linear relation of the swimmer-type para-
meter wand the force-dipole coefficient wBA
2
[Fig. 1(b)]. This
is expected since the terminal values of the envelope function
f(x) in eqn (2), which determine the strength of the force
dipole, are linear in w. Furthermore, we find that due to the
definition of the envelope function with hf(x)i= 1, the swim-
ming velocity is nearly independent of w[cf. Fig. 1(b), red curve].
3.2 Force-dipole flow fields in the Hele-Shaw geometry
As already explained, our study of the collective dynamics of the
squirmer rods is performed in a Hele-Shaw cell, which alters
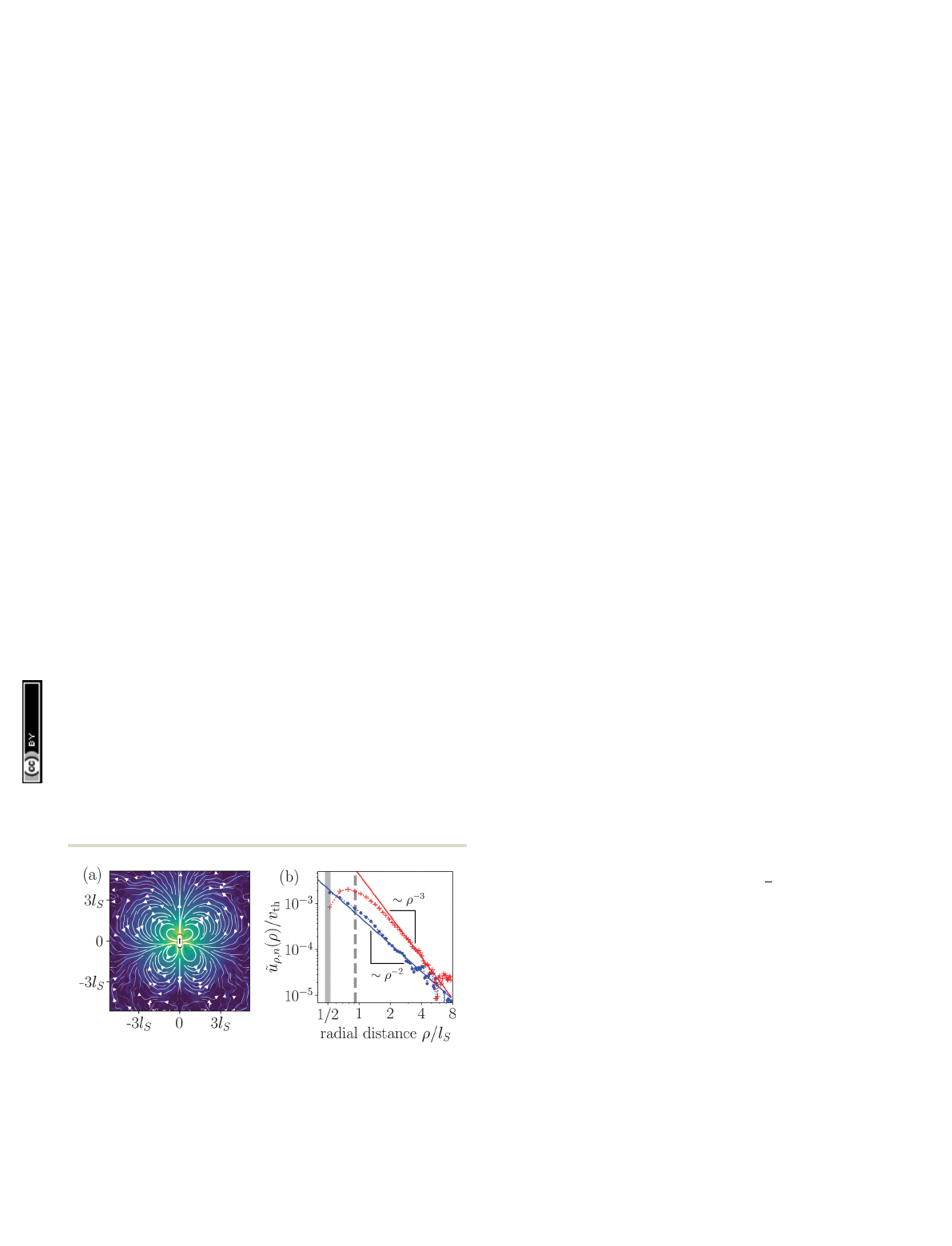
Fig. 2 Hydrodynamic flow field around a pusher-type squirmer rod
(w=1.0) swimming in the Hele-Shaw geometry with cell height Dz=6R.
(a) Flow field in the mid plane of the Hele-Shaw cell. The force dipole clearly
dominates the near field. (b) Radial components u
˜
r,n
(r)oftheleadingsource-
dipole (blue) and force-dipole (red) flow fields. They are normalized by the
thermal velocity of fluid particles, vth ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
kBT=m0
p.
Paper Soft Matter
Open Access Article. Published on 05 July 2022. Downloaded on 10/27/2022 10:58:01 AM.
This article is licensed under a
Creative Commons Attribution 3.0 Unported Licence.
View Article Online

This journal is © The Royal Society of Chemistry 2022 Soft Matter, 2022, 18, 6179–6191 | 6183
the radial decay of the multipole far fields but also their
strengths depends on the cell height Dz.
76,99
Thus, the multi-
pole expansion for the radial component of the flow field
becomes ~
urðr;jÞ¼P
1
n¼1ðAnþBnÞrðnþ1ÞcosðnjÞfor a micro-
swimmer oriented along the x-axis.
76
Force multipoles dissipate
momentum at the bounding walls, which increases for smaller
Dz,i.e., when they are closer to the walls.
99,108
In our notation
this means that the coefficient of the force dipole in the
Hele-Shaw cell scales as A
2
BA
2
Dzwith respect to the bulk
coefficient A
2
. In contrast, the fluid mass flux initiated by a
point source in the Hele-Shaw geometry is distributed in a
volume that is proportional to the cell height Dz. Hence, the
coefficient of the source dipole, as all the other source multi-
poles, scales as B
2
BB
2
/Dz.
76,99
Fig. 2(a) shows the flow field of a pusher-type squirmer rod
(w=1.0, a= 3.25) swimming in Hele-Shaw geometry with wall
distance Dz=6R. The force-dipole field visibly dominates the
flow field with its characteristic outwards directed streamlines
along the rod and inwards directed streamlines at the side.
From the simulated flow field, we extracted the radial parts
of different hydrodynamic multipoles u
˜
r,n
(r) following our
previous work,
76
and arrived at the curves shown in Fig. 2(b).
The radial part of the force dipole Br
3
(red symbols) domi-
nates the flow field up to a distance of circa 8l
S
, where it is
exceeded by the more long-ranged field of the source dipole
Br
2
(blue symbols). For smaller cell heights Dz, this crossover
occurs at smaller distances. Since we are interested in exploring
the effect of the hydrodynamic force dipole, we keep Dz=6Rfor
the rest of this work, such that the force dipole dominates the
flow field close to the squirmer rod.
3.3 Pushers-type squirmer rods
3.3.1 State diagram. For the minimum dipole strength
w=1, i.e., the pusher-type squirmer rods with the largest
force dipole, we study the collective dynamical states as a
function of the aspect ratio aand area fraction f.Acomparison
between the state diagrams of neutral squirmer rods [cf. Fig. 1(c)]
and pusher rods [cf. Fig. 3(a)] already illustrates the strong
influence of the long-ranged hydrodynamic dipole–dipole inter-
actions. Compared to neutral squirmer rods the transition line
from the disordered to other states is shifted to larger area
fractions and new dynamic states arise such as active turbulence
and dynamic clustering. We provide an overview of the observed
dynamic states before presenting a more quantitative analysis in
Section 3.3.2.
Most interestingly, compared to neutral squirmer rods the
flow field of the pusher rod obviously suppresses the formation
of dynamic swarms and impedes the single swarm state, which
only occurs for large aZ4.0. This is in agreement with findings
of Saintillan and Shelley,
88
who employ slender-body theory to
show that polar and nematic order in systems of elongated
pusher microswimmers is destroyed by their hydrodynamic
flow fields. Likewise, explicit hydrodynamic simulations of
collective dynamics of spherical squirmers show that pushers
create disordered homogeneous systems,
13,109
while pullers
show swarming.
109
An argument for this difference in the
collective dynamic behavior is found in both implicit
90
and
explicit
71
hydrodynamic simulations, which show that pushers
are deflected during collision, while pullers align.
Instead of the suppressed swarming states for ar3.25,
we find a very dynamic or turbulent state. A typical snapshot of
a rod configuration is shown in Fig. 4(a), a video is provided
in the Video 1 (ESI†). Here, the hydrodynamic dipole–dipole
interactions compete with steric interactions that favor the
formation of single swarms at large area fraction fin the case
of neutral rods.
94
Towards lower fthe turbulent state transi-
tions to the disordered state and towards higher aspect ratio a
to cluster or swarm states.
Similar to other examples of active turbulence in theory and
experiments,
47,49–51,97
we find two cascades in the power spec-
tral density of the squirmer velocities, as we will show in detail
in Section 3.3.2. For the turbulent state at a= 3.25 and f= 0.6
we construct a continuous velocity field v(x,y) to visualize the
turbulent flow pattern and its vortices. A snapshot of the system
and the resulting flow field are shown in Fig. 4(a) and (b),
respectively. We also calculate the vorticity o=(rv)
z
, which
is shown in Fig. 4(c). To easily obey the periodic boundary
conditions, the vorticity was determined via a Fourier trans-
formation. In all turbulent states, we observe that squirmer
rods show a local alignment, which extends over short streaks,
where squirmer rods swim side by side and head to tail.
However, these streaks buckle and dissolve frequently, leaving
a chaotic pattern of streaks and vortices.
Fig. 3 (a) State diagram of the strongest pusher-type squirmer rods with
w=1 in the parameter space aspect ratio aversus area fraction f.
(b) Snapshot of a single swarm state at a= 4.75 and f= 0.6, (c) snapshot of
a dynamic cluster state at a= 4.0 and f= 0.7 as indicated in the state
diagram. The color of the individual squirmer rods encode their orientation
e
ˆ
i
in the xy-plane.
Soft Matter Paper
Open Access Article. Published on 05 July 2022. Downloaded on 10/27/2022 10:58:01 AM.
This article is licensed under a
Creative Commons Attribution 3.0 Unported Licence.
View Article Online
Loading more pages...