
ARTICLE
Molecular basis of antibiotic self-resistance in a
bee larvae pathogen
Tam Dang 1, Bernhard Loll 2, Sebastian Müller1, Ranko Skobalj1, Julia Ebeling 3, Timur Bulatov1,
Sebastian Gensel1, Josefine Göbel 3, Markus C. Wahl2,4, Elke Genersch3,5, Andi Mainz1&
Roderich D. Süssmuth 1✉
Paenibacillus larvae, the causative agent of the devastating honey-bee disease American
Foulbrood, produces the cationic polyketide-peptide hybrid paenilamicin that displays anti-
bacterial and antifungal activity. Its biosynthetic gene cluster contains a gene coding for the
N-acetyltransferase PamZ. We show that PamZ acts as self-resistance factor in Paenibacillus
larvae by deactivation of paenilamicin. Using tandem mass spectrometry, nuclear magnetic
resonance spectroscopy and synthetic diastereomers, we identified the N-terminal amino
group of the agmatinamic acid as the N-acetylation site. These findings highlight the phar-
macophore region of paenilamicin, which we very recently identified as a ribosome inhibitor.
Here, we further determined the crystal structure of PamZ:acetyl-CoA complex at 1.34 Å
resolution. An unusual tandem-domain architecture provides a well-defined substrate-bind-
ing groove decorated with negatively-charged residues to specifically attract the cationic
paenilamicin. Our results will help to understand the mode of action of paenilamicin and its
role in pathogenicity of Paenibacillus larvae to fight American Foulbrood.
https://doi.org/10.1038/s41467-022-29829-w OPEN
1Institut für Chemie, Technische Universität Berlin, Berlin, Germany. 2Institut für Chemie und Biochemie, Strukturbiochemie, Freie Universität Berlin,
Berlin, Germany. 3Institute for Bee Research, Department of Molecular Microbiology and Bee Diseases, Hohen Neuendorf, Germany. 4Macromolecular
Crystallography, Helmholtz Zentrum Berlin für Materialien und Energie, Berlin, Germany. 5Institut für Mikrobiologie und Tierseuchen, Fachbereich
NATURE COMMUNICATIONS | (2022) 13:2349 | https://doi.org/10.1038/s41467-022-29829-w |www.nature.com/naturecommunications 1
1234567890():,;
Pollination of wild and cultivated flowering plants is an
indispensable ecosystem service, which is mainly provided
by pollinating insects. Among the insect pollinators, man-
aged honey bee colonies play a particularly important role in
agriculture, where they are widely used as commercial pollinators
and contribute to 35% of the production volume of global food
crops1. In order to secure human food supply, it is therefore
important to ensure the health of honey bees, which is con-
tinuously threatened by the overuse of insecticides such as
neonicotinoid2in agriculture and also by various viral, bacterial,
and fungal pathogens as well as metazoan parasites3.
The Gram-positive, facultative anaerobic, spore-forming bac-
terium, Paenibacillus larvae (P. larvae), is the causative agent of
the epizootic American Foulbrood (AFB) of honey bees4. AFB is
the most serious bacterial disease of honey bees and is classified as
notifiable disease in most countries because it is highly contagious
and lethal to entire colonies. Furthermore, most authorities
consider the killing of diseased colonies and burning of the hive
material the only workable control measure resulting in con-
siderable economic losses in apiculture. AFB is a fatal intestinal
infection of the honey bee brood initiated in first instar larvae by
ingestion of spore-contaminated food. The distribution of the
spores, the infectious form of P. larvae, within a colony and
between colonies, also within apiary and between apiaries5,
consequently leads to honey bee colony losses. The use of
enterobacterial repetitive intergenic consensus (ERIC) sequence
primers has revealed four well-described genotypes ERIC I to
ERIC IV4for P. larvae which differ in virulence on the larval6and
colony level7as well as in pathogenesis strategies employed to kill
the host8. The existence of another ERIC genotype, ERIC V, has
recently been proposed9. From contemporary outbreaks of AFB
all over the world, only P. larvae ERIC I and ERIC II can be
isolated10, suggesting that the hypervirulent genotypes ERIC III
to ERIC V did not become established in the honey bee
population.
In our quest to find sustainable control measures against this
most serious bacterial disease of honey bees, we started to unravel
AFB pathogenesis by analyzing the interaction between P. larvae
and honey bee larvae on a molecular level. We identified several
virulence factors of P. larvae ERIC I and ERIC II and showed that
two AB toxins11,12, a chitin-degrading enzyme13,14 and also an
S-layer protein15,16 have a pivotal role in the virulence of this
pathogen and that P. larvae also produces various secondary
metabolites17. Bacterial secondary metabolites, with polyketides
and (non-)ribosomal peptides as important representatives, pro-
vide highly valuable lead structures, among them antibiotics with
novel modes of action for drug development to fight various
infectious diseases18,19. Secondary metabolites can also act as
virulence(-like) factors, functioning as signal molecules in gene
regulation of defense or growth mechanisms20–22. The search for
secondary metabolites produced by P. larvae led to the structural
elucidation of paenilamicin that shows cytotoxic, antibacterial
and antifungal activities23,24. It is currently assumed that paeni-
lamicin is produced as a defense molecule against microbial
competitors, since only P. larvae can usually be isolated as a pure
culture from the cadavers of AFB-killed larvae, suggesting that
other saprophytic competitors are absent in the degradation
process of the larval cadavers to the characteristic ropy mass25.
We recently substantiated this view by showing that paenilamicin
was active against the bee-associated saprophyte P. alvei in
P. larvae-infected larvae23.
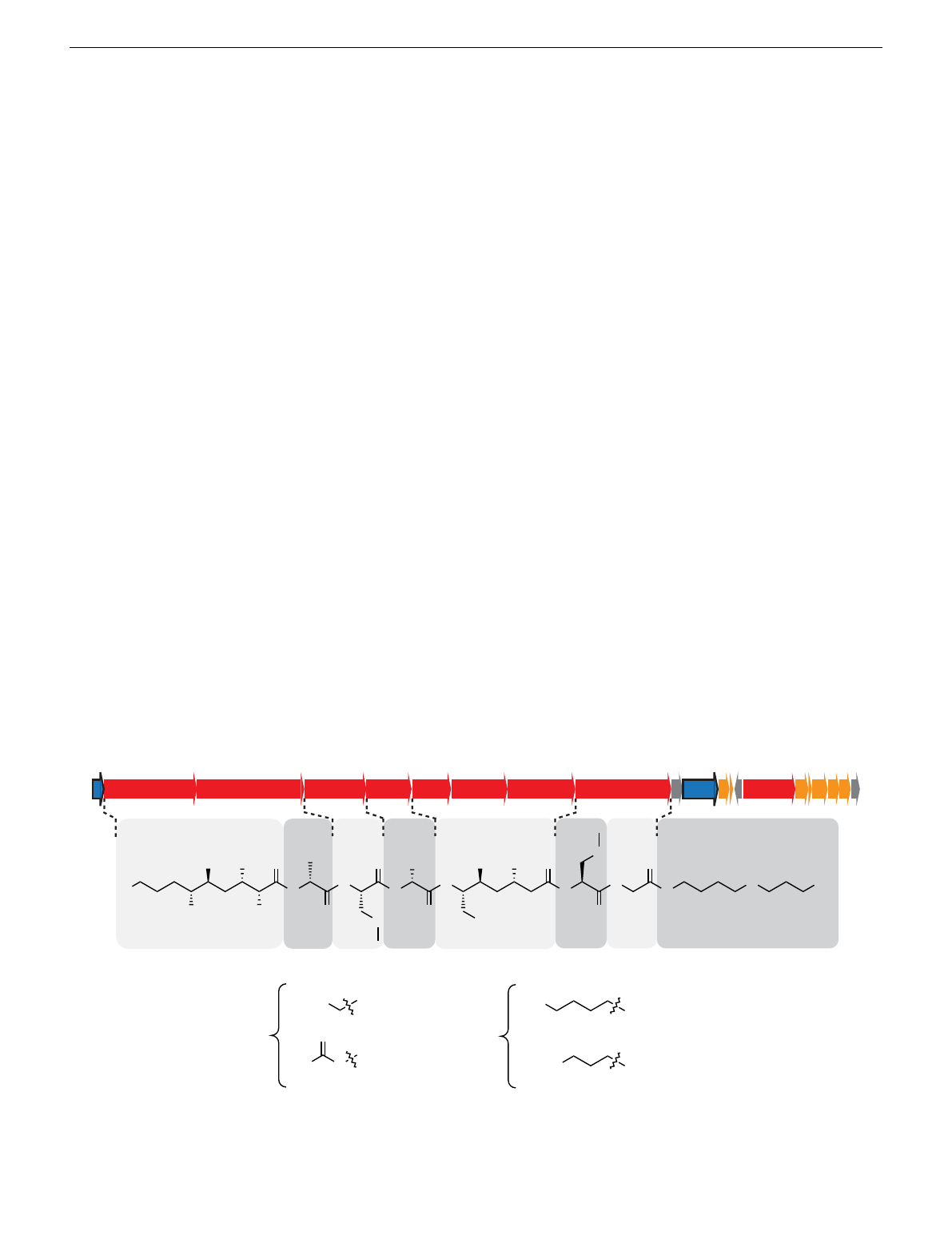
Paenilamicin is a linear, cationic aminopolyol peptide anti-
biotic and is synthesized via an unusual nonribosomal peptide
synthetase-polyketide synthase (NRPS-PKS) hybrid assembly line
that exhibits several fascinating biosynthetic features. It contains
unusual structural motifs such as galantinamic acid (Glm),
agmatinamic acid (Aga), N-methyldiaminopropionic acid
(mDap), galantinic acid (Gla) and a 4,3-spermidine (Spd) at the
C-terminus (Fig. 1). P. larvae produces a mixture of paenilamicin
variants A1, A2, B1, and B2. They only differ in two positions of
the paenilamicin backbone: at the N-terminus and in the center
between mDap1 and Gla. Either a lysine (series A) or an arginine
(series B) is activated by the adenylation domain of NRPS1
(Fig. 1). The amino acid residue between mDap1 and Gla is a
lysine (series 1) or an ornithine (series 2) assigned to be incor-
porated by NRPS4 (pamD), respectively (Fig. 1).
The pam gene cluster harbors a gene encoding the putative
acetyl-CoA-dependent N-acetyltransferase PamZ, which belongs
to the Gcn5-related N-acetyltransferase (GNAT) superfamily26,27.
One prominent member of this superfamily is the bacterial
(Lys, series 1)
(Lys, series 1)
(Orn, series 2)
(Orn, series 2)
(Glm, series A)
(Glm, series A)
(Aga, series B)
(Aga, series B)
H2N
N
H
H2N
NH
H2N
H2N
R1 =R
2 =
Ala mDap1 mDap2Lys/OrnGlm/Aga Gla Gly Spd
ZpamA pamB pamC pamD pamE pamF pamG pamH I JK pamNMLOSRQP
NRPS1-PKS1 PKS2-NPRS2 NRPS3 NRPS4 NRPS5 PKS3 PKS4 NRPS6-NRPS7 NRPS0
N
H
O
OH
OHOH
NH2
R1
O
H
N
O
N
H
NH
R2
H
N
OOH
OH OH
N
H
ONH
H
N
O
N
H
OH
NNH2
Fig. 1 Biosynthetic gene cluster and structure of paenilamicin variants. The paenilamicin (pam) gene cluster23 contains core biosynthetic (red), auxiliary
biosynthetic (orange), resistance (pamZ and pamJ; blue) and other (gray) genes and expresses the nonribosomal peptide synthetase-polyketide synthase
(NRPS-PKS) hybrid biosynthetic machinery for the production of paenilamicin A1 (Glm, Lys), A2 (Glm, Orn), B1 (Aga, Lys), and B2 (Aga, Orn). Glm -
galantinamic acid, Aga - agmatinamic acid, Lys lysine, Orn - ornithine, Ala - alanine, mDap - N-methyldiaminopropionic acid, Gla - galantinic acid, Gly -
glycine, Spd - 4,3-spermidine.
ARTICLE NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-022-29829-w
2NATURE COMMUNICATIONS | (2022) 13:2349 | https://doi.org/10.1038/s41467-022-29829-w | www.nature.com/naturecommunications

aminoglycoside N-acetyltransferase (AAC) that plays an impor-
tant role in antibiotic resistances, particularly in clinical and
environmental settings28. Aminoglycoside antibiotics have been
widely used in the treatment of bacterial infections but they
rapidly lose activity against multi-resistant bacteria due to
adaptation and the development of resistance. By contrast, self-
resistance is an innate, non-adaptation-based mechanism for the
protection against self-produced antimicrobial agents. Since self-
produced antimicrobial agents could also harm the bacterial host,
self-resistance is critical for survival and territorial competition.
Our results demonstrate the deactivation of paenilamicins by
the regio- and stereoselective self-resistance protein PamZ
including its high-resolution crystal structure that shows how its
tandem-domain arrangement may organize substrate binding.
Together with a parallel study29, in which we report on the total
synthesis and the biological evaluation of paenilamicin, we have
here unambiguously identified the N-terminal building block of
paenilamicins as an essential switch for target binding, biological
activity and self-resistance.
Results and discussion
Regio- and stereoselective N-acetylation of paenilamicin by
PamZ. To confirm our hypothesis that PamZ (NCBI WP_
023484187) is an acetyl-CoA-dependent N-acetyltransferase that
targets paenilamicins, we monitored PamZ-mediated anti-
bacterial effects in vitro by agar diffusion assays against Bacillus
megaterium (B. megaterium) as indicator strain as well as by mass
spectrometry (MS) and nuclear magnetic resonance (NMR)
spectroscopy. To this end, the pamZ gene was amplified from the
wild-type (WT) P. larvae ERIC II strain, inserted into the com-
mercial pET28a(+) vector, and transformed into E. coli BL21-
Gold(DE3) for heterologous expression. PamZ was then purified
(Supplementary Fig. 1) and used for the assays including four
native paenilamicin variants as substrates and acetyl-CoA as co-
substrate. The paenilamicin variants were purified from P. larvae
ERIC I and ERIC II, which preferably produce the paenilamicin
mixtures A2/B2 and A1/B1, respectively (Fig. 1and Supple-
mentary Fig. 2). In addition, we also tested synthetic paenilamicin
B2 (PamB2_3)29.
The agar diffusion assays clearly showed that paenilamicins
incubated with PamZ and acetyl-CoA were not able to inhibit the
growth of B. megaterium, whereas antibacterial activity was
observed in the absence of acetyl-CoA and/or PamZ (Fig. 2). This
loss of biological activity correlated with the conversion of
paenilamicins to the corresponding N-acetylpaenilamicins as
observed by HPLC-ESI-MS. ESI mass spectra revealed that the
mass-to-charge ratios of natural and synthetic paenilamicins
exhibited a characteristic mass shift of 42 Da indicative of the
addition of an acetyl group (Supplementary Figs. 3–7).
Paenilamicin contains several primary and secondary amino
groups that are potential candidates for N-acetylation. To determine
the site of acetylation, we monitored PamZ-mediated effects in
fingerprint tandem MS and NMR spectra of paenilamicin before
and after treatment with PamZ/acetyl-CoA. Besides the mass shift of
42 Da for the acetylation, characteristic MS2fragmentation patterns
originated from the difference between Glm and Aga residues in
series A and B (+28 Da) as well as the difference between Lys and
Ornresiduesinseries1and2(+14 Da). MS2fragmentation mainly
resulted in fragment ions b
4
,y
4
and y
6
of each paenilamicin and N-
acetylpaenilamicin variant acquired by collision-induced dissociation
(Supplementary Table 1). Fragment ion b
4
varied depending on the
paenilamicin series showing mass shifts of 14 Da and 28 Da.
Importantly, we observed a mass shift of 42 Da only for fragment
ion b
4
, indicating acetylation in the N-terminal half of paenilamicin.
By contrast, the fragment ions y
4
and y
6
did not exhibit any mass
shifts of 42 Da between paenilamicins and N-acetylpaenilamicins.
Thus, we excluded acetylation in the C-terminal half of paenilamicin
(Supplementary Figs. 8–18). In addition, we detected and isolated
small amounts of N-acetylpaenilamicin A1, B1, and B2 from
supernatants of P. larvae ERIC I and ERIC II (Supplementary
Fig. 19), and compared them with our products formed in vitro. The
MS2fragmentation analysis confirmed that the mono-acetylation in
the N-terminal half of paenilamicin also occurred in vivo
(Supplementary Figs. 20–22). The MS2experiments did not reveal
whether the N-terminal amino group of Aga/Glm or its side chain
(amino/guanidino group) was acetylated.
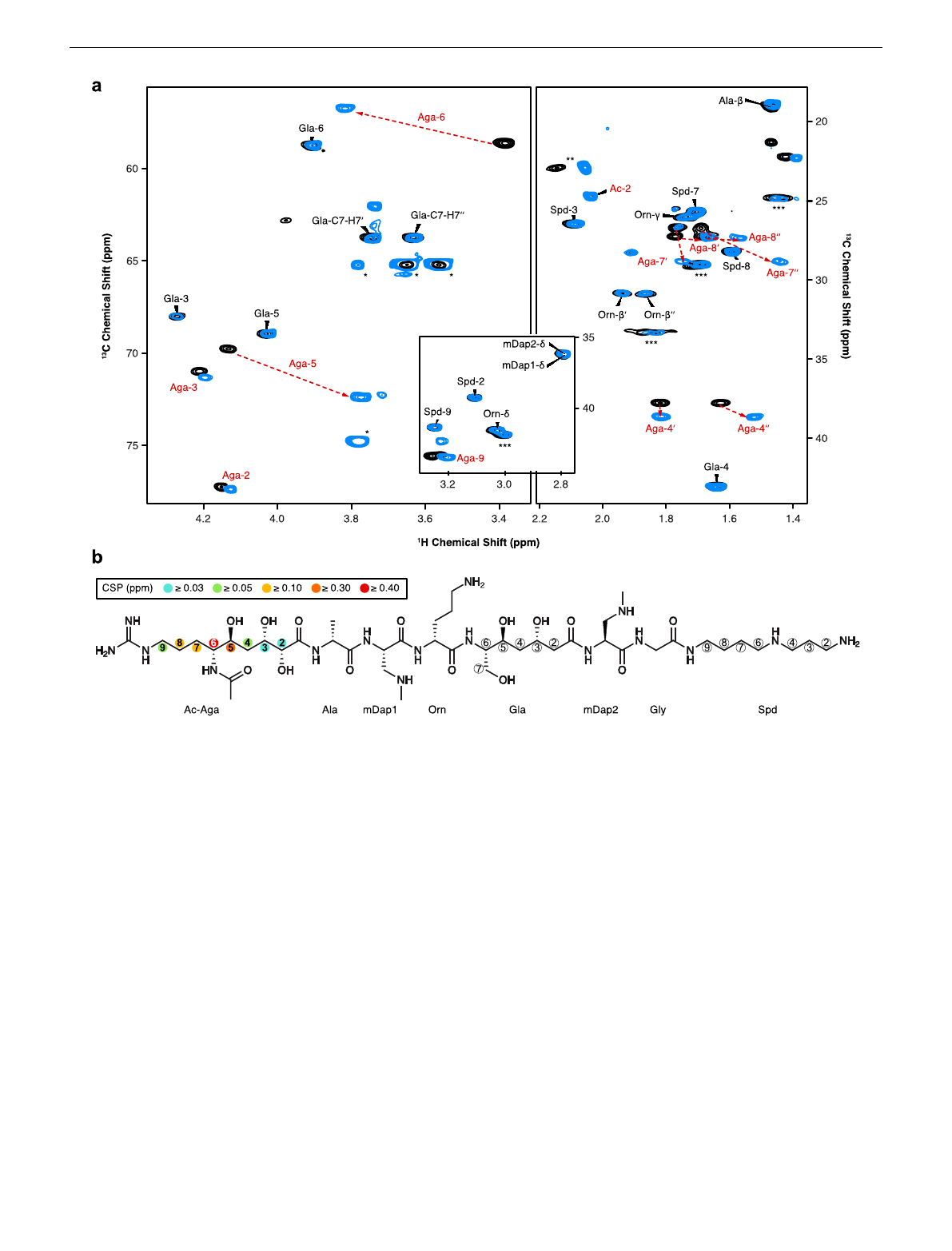
To ultimately identify the functional group that is modified by
PamZ, we acquired 1H-13C hetero-nuclear single-quantum
coherence (HSQC) NMR spectra of paenilamicin B2 before and
after incubation with PamZ/acetyl-CoA. Although both spectra
were mostly superimposable, severe chemical shift perturbations
(CSPs) were observed for a minor fraction of cross-peaks (Fig. 3a).
Mapping CSPs onto the structure of paenilamicin B2 revealed a
well-defined region comprising the N-terminal half, with the
strongest effect being located at position 6 of Aga (Fig. 3b and
Supplementary Table 2). N-acetylpaenilamicin B2 also showed an
additional cross-peak compared to paenilamicin B2, which we
tentatively assigned to the methyl moiety of the newly attached
acetyl group (Fig. 3a). Our data unequivocally demonstrated that
PamZ mono-N-acetylates the N-terminal amino group at Aga-6
position of paenilamicin and thereby abolishes its antibacterial
activity. Ultimately, this result is further supported by two
synthetic diastereomers of paenilamicin B2 with L- instead of the
native D-configuration at Aga-6 (PamB2_1 and PamB2_2), that
were both antibacterially less active29 and that were not modified
by PamZ (Fig. 4and Supplementary Fig. 23).
The structure of PamZ:acetyl-CoA binary complex. A BLAST30
search indicated that PamZ belongs to the GNAT superfamily
with a sequence identity of 31% to the N-acetyltransferase, ZmaR,
whose structure has not yet been determined and which confers
resistance against the aminopolyol peptide antibiotic, zwittermi-
cin A, in Bacillus cereus UW85 (Supplementary Fig. 24)31.We
solved the crystal structure of PamZ in complex with acetyl-CoA
at a resolution of 1.34 Å by using the uncharacterized N-acetyl-
transferase from Streptococcus suis 89/1591 (PDB 3G3S) for
molecular replacement (Supplementary Table 3). The electron
density was of excellent quality, allowed the modeling of the
entire polypeptide chain, and unambiguously revealed the bound
acetyl-CoA (Supplementary Fig. 25). PamZ comprises an
N-terminal domain (NTD, residues 1–128, secondary structure
elements indicated by primes) and a C-terminal domain (CTD,
Fig. 2 Deactivation of paenilamicins through PamZ-mediated N-
acetylation tested by agar diffusion assay against B. megaterium as the
indicator strain. Paenilamicin variants (PamA1, A2, B1, B2) isolated from P.
larvae and synthetic paenilamicin B2 (PamB2_3) were incubated in vitro
with both acetyl-CoA and PamZ (1), acetyl-CoA only (2), or PamZ only (3).
Samples 2 and 3 are negative controls and indicate the lack of bacterial
growth.
NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-022-29829-w ARTICLE
NATURE COMMUNICATIONS | (2022) 13:2349 | https://doi.org/10.1038/s41467-022-29829-w |www.nature.com/naturecommunications 3
residues 140–275), which both adopt the characteristic GNAT
fold (Fig. 5a)32. The two tandem-GNAT domains, that may have
originated from a gene duplication event, share low sequence
identity (<20%) and are connected by an α-helical linker (α
bridge
,
residues 129–139). The overall fold of each domain is very similar
to that of bacterial aminoglycoside N-acetyltransferases (AACs),
as pairwise structural alignments with several AACs (PDB 1BO4,
1M4I,1S3Z) gave root-mean-square deviations (RMSDs) of
2.9–4.2 Å for both the NTD and CTD (Supplementary Fig. 26)33.
A structural superposition of the NTD and CTD of PamZ yielded
an RMSD of 4.2 Å for 75 pairs of C
α
atoms (Supplementary
Fig. 27)33.
However, a comparison with the typical GNAT fold revealed
several unique features in PamZ. Instead of two N-terminal α-
helices, α1 and α2, both domains of PamZ contain three short
helical segments, α0-α1-α2(α0’–α1’–α2’), which pack onto one
face of the central antiparallel β-sheet, β2–β3–β4(β2’–β3’–β4’),
whereas helix α3(α3’) buries its other side. A kink in the
backbone conformation of strand β3, involving residues T199 and
C200, causes a strong right twist and thus a distortion of the
antiparallel β3–β4 arrangement, which led us to discriminate
these strands as β3a/β3b and β4a/β4b (Fig. 5a). The central β-
sheet is extended by strand β5’in the NTD, whereas the CTD
shows the characteristic β-bulge of GNAT enzymes —a
V-shaped cavity between strands β4b and β5 accommodates the
pantetheine segment of CoA (Fig. 6a). Furthermore, the well-
conserved pyrophosphate-binding loop (P-loop) of the GNAT
family (R/Q-X-X-G-X-A/G)26 is only present in the CTD of
PamZ (Q-N-K-G-L-A) between strand β4b and helix α3
(Fig. 6a)34, whereas the NTD is missing this signature motif.
Accordingly, there is only one acetyl-CoA molecule canonically
bound in the PamZ structure, namely to the CTD.
Hence, we concluded that the NTD is incompetent in binding
acetyl-CoA and rather plays a structural role, in particular for
substrate binding (see below). Notably, many GNAT enzymes exist
as homodimers in solution with various arrangements of the
monomer-monomer interface32. Likewise, AACs have often been
crystallographically observed in a homodimeric state, although
their quaternary structure in solution may vary35.PamZexistsasa
monomer, both in solution and in the crystal (Supplementary
Fig. 3 Identification of the N-acetylation site through 2D NMR spectroscopy. a Overlay of relevant 1H-13C HSQC sections of paenilamicin B2 (black) and
N-acetylpaenilamicin B2 (blue). Strongly perturbed cross-peaks are highlighted with red labels. Known impurities are labeled with one, two and three
asterisks arising from glycerol, acetic acid and residual purification traces of paenilamicin B1, respectively. The numbers (in backbone) and Greek letters (in
amino acid residues) refer to atom positions in the corresponding building blocks as labeled in b.bSignificant chemical shift perturbations (CSPs) and
corresponding positions are indicated as circles in the chemical structure of N-acetylpaenilamicin B2 (see legend for color code). Aga - agmatinamic acid,
Ala - alanine, mDap - N-methyldiaminopropionic acid, Orn - ornithine, Gla - galantinic acid, Gly - glycine, Spd - 4,3-spermidine, Ac - acetyl group.
ARTICLE NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-022-29829-w
4NATURE COMMUNICATIONS | (2022) 13:2349 | https://doi.org/10.1038/s41467-022-29829-w | www.nature.com/naturecommunications
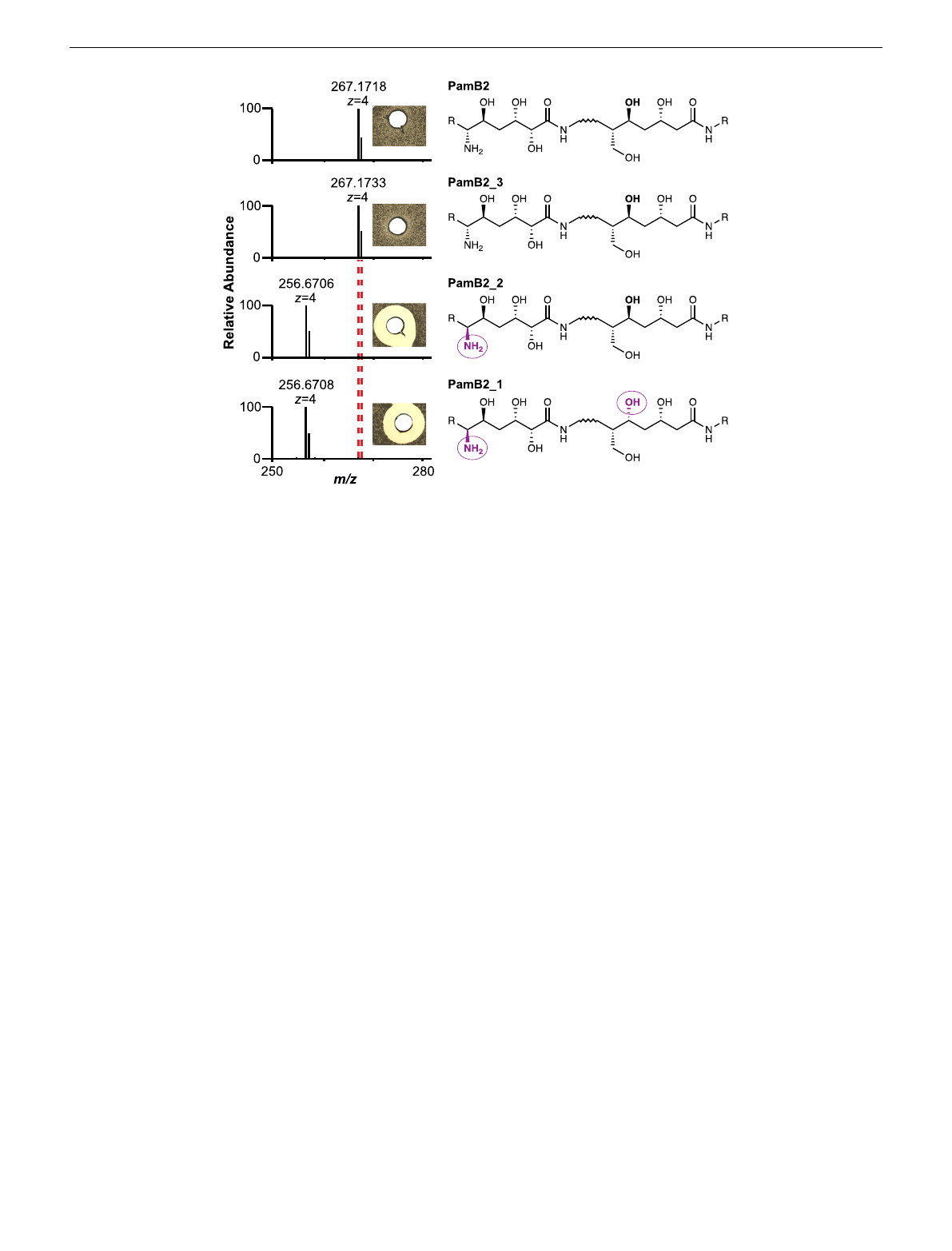
Fig. 28). However, the tandem-GNAT domain constellation of
PamZ achieves an intramolecular domain-domain interface that
resembles that of some GNAT homodimers. There are several
GNAT enzymes that utilize domain swapping of strand β6to
stabilize their homodimeric structure34,36,37. Interestingly, a major
interface in PamZ is achieved by domain swapping of strand β6
(β6’), which inserts between strands β5’and β6’(β5andβ6) of the
opposing domain and thus forms an extended, antiparallel, and
strongly-twisted β-sheet throughout the enzyme (Fig. 5b). This β-
sheet is only interrupted by the β-bulge in the CTD accommodat-
ing the cofactor and allowing the amide groups of its pantetheine
portion to form pseudo-β-sheet hydrogen bonds to strand β4b
(Fig. 6a). A very similar tandem arrangement of a pseudo-GNAT
NTD and a canonical GNAT CTD can be found in the template
protein (PDB 3G3S). Another example is the structure of
mycothiol synthase MshD from Mycobacterium tuberculosis,
which is also organized as a tandem repeat of two GNAT domains
with a catalytically inactive NTD38.
PamZ appears to utilize its NTD to form a well-defined
substrate pocket with strands β5 and β6’representing its floor. A
second interface between the NTD and CTD is accomplished
through tight packing of helix α2’onto the small β3b–β4a sheet.
Further interactions involve helix α2 of the CTD and the loops
between α2’and β2’as well as β3’and β4’of the NTD. These
inter-domain contacts fully cover the central groove that is
normally found at the interface of homodimeric structures of
GNAT enzymes and restrict substrate entry to the opening that is
also used by the cofactor. This remaining cleft between the two
domains of PamZ is decorated with several acidic residues (e.g.
E89, E116, E118, D120, D162, D170, D215, E216, E217, E218,
E272, E274, and the C-terminus) and thus deploys a large
negatively charged surface to attract its polycationic substrate
(Fig. 5c). A corridor that lies aside and beyond the acetyl group of
the cofactor is approximately 7–8 Å deep and 8–9 Å wide with
respect to the thioester carbonyl atom. Although we did not
obtain crystals of a ternary PamZ-acetyl-CoA-paenilamicin
complex, the position of acetyl-CoA, the well-defined shape of
the neighboring pocket and our knowledge about the substrate’s
N-terminal acetylation site allows us to predict that the Glm/Aga
side chain of paenilamicin very likely penetrates into this pocket.
Acidic residues D25 (loop between α1’and α2’), E122 (β6’), and
E208 (β4a) are well-positioned within the pocket to accommodate
and stabilize the guanidine group of Aga, as well as to tolerate the
Nζamine of Glm. Other residues that shape the substrate pocket
include T58/T59 (loop between β3’and β4’), T98 (β5’) and Y124
(β6’) of the NTD as well as C200/Y201 (β3b) and S245/F247 (β5)
of the CTD (Fig. 6c). This shows that both domains most likely
contribute to substrate recognition. Moreover, the structure of
PamZ explains its regioselectivity: if PamZ was to modify e.g. the
terminal amino group of spermidine in paenilamicin, the enzyme
would not require such a deep substrate-binding pocket. The
architecture of the central groove between the NTD and CTD has
evolved to optimally accommodate the N-terminal Glm/Aga
building block of paenilamicin, whilst terminal amines such as
those of spermidine, ornithine and lysine side chains would not
occupy this binding pocket, as they would experience significantly
less binding stabilization.
Such accommodation of Glm/Aga in the substrate pocket
would position the N-terminal amino group of Aga-6 close to the
thioester carbonyl of the cofactor. An active site aspartate or
glutamate residue commonly acts as a general base to trigger the
N-acetylation reaction by deprotonation of the amine followed by
a nucleophilic attack at the carbonyl of the thioester35. In PamZ,
the side chains of E122 (β6’) as well as E208 (β4a) exhibit an
interatomic distance of ~7 Å to the carbonyl atom of acetyl-CoA
and thus might be in close proximity to the N-terminal amino
group of Aga-6 (Fig. 6c). Residue S245 (β5) is sandwiched
between E122 and E208, and may mediate deprotonation and/or
proton shuttling. Furthermore, we cannot exclude the involve-
ment of water molecules during proton transfer. An oxyanion
Fig. 4 Substrate specificity and stereoselectivity of PamZ. The natural product (PamB2), synthetic paenilamicin B2 (PamB2_3) and synthetic
diastereomers of paenilamicin B2 (PamB2_2,PamB2_1) were incubated with PamZ and acetyl-CoA in vitro and tested in an agar diffusion assay against
Bacillus megaterium (insets). From the chemical structure of paenilamicins, only the agmatinamic and galantinic acid are depicted to emphasize the changes
in stereoconfiguration highlighted in purple and circles. Each single reaction was verified by HPLC-ESI-MS. Dashed lines indicate the mass shift of 42 Da
(4 × 10.5 Da) due to N-acetylation.
NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-022-29829-w ARTICLE
NATURE COMMUNICATIONS | (2022) 13:2349 | https://doi.org/10.1038/s41467-022-29829-w |www.nature.com/naturecommunications 5
Loading more pages...