
nutrients
Review
Zinc and Sepsis
Wiebke Alker 1,2 and Hajo Haase 1,2,*ID
1Department of Food Chemistry and Toxicology, Berlin Institute of Technology, 13355 Berlin, Germany;
2
TraceAge—DFG Research Unit on Interactions of Essential Trace Elements in Healthy and Diseased Elderly,
Potsdam-Berlin-Jena, Germany
*Correspondence: [email protected]; Tel.: +49-30-314-727-01
Received: 25 June 2018; Accepted: 24 July 2018; Published: 27 July 2018
Abstract:
Sepsis, defined as a “life-threatening organ dysfunction caused by a dysregulated
host-response to infection” is a major health issue worldwide and still lacks a fully elucidated
pathobiology and uniform diagnostic tests. The trace element zinc is known to be crucial to ensure an
appropriate immune response. During sepsis a redistribution of zinc from serum into the liver has
been observed and several studies imply a correlation between zinc and sepsis outcome. Therefore the
alterations of zinc concentrations in different tissues might serve as one part of the host’s defense
mechanism against pathogens during sepsis by diverse mechanisms. It has been suggested that zinc
is involved in nutritional immunity, acts as a hepatoprotective agent, or a differentiation signal for
innate immune cells, or supports the synthesis of acute phase proteins. Further knowledge about
these events could help in the evaluation of how zinc could be optimally applied to improve treatment
of septic patients. Moreover, the changes in zinc homeostasis are substantial and correlate with the
severity of the disease, suggesting that zinc might also be useful as a diagnostic marker for evaluating
the severity and predicting the outcome of sepsis.
Keywords: zinc; sepsis; biomarker; supplementation; homeostasis
1. Introduction
Zinc is of fundamental importance for the immune system and is involved in different pathologies.
In recent years, indications have appeared that zinc homeostasis might be an important factor
during sepsis. The following review focuses on the alterations of zinc homeostasis during sepsis
and possible physiological functions of this process. It further discusses potential risks and benefits of
zinc supplementation as well as a possible approach for using serum zinc as a biomarker for sepsis.
1.1. Sepsis
The term “sepsis” in relation to a disease has already been used by Hippocrates, but to this day it
remains a challenge to compile a definition comprising its complexity [
1
]. This results from the fact
that sepsis is rather a syndrome than an illness, showing a not yet fully elucidated pathobiology, and
with uniform diagnostic tests still lacking [
2
]. Sepsis is responsible for about 6 million deaths per year,
making it a critical illness and one of the major causes of mortality worldwide [
3
,
4
]. Its epidemiological
burden is assumed to be much higher in low- and middle-income countries and the mortality rate is
affected by the global national income [3,5].
There is an urgent need for an easily understandable definition in order to establish public
awareness, as well as for improved and uniform diagnostic guidelines for an early recognition of
sepsis [
2
,
3
]. In the past, different task forces have approached these issues [
6
,
7
]. A recent consensus
defined sepsis as a “life-threatening organ dysfunction caused by a dysregulated host-response to
Nutrients 2018,10, 976; doi:10.3390/nu10080976 www.mdpi.com/journal/nutrients

Nutrients 2018,10, 976 2 of 17
infection” (Sepsis-3) [
2
]. To diagnose organ dysfunction in the clinical setting, Singer et al. recommend
the Sequential Organ Failure Assessment (SOFA) score. It includes parameters to evaluate the functions
of respiration, the liver, the cardiovascular system, the central nervous system, the kidneys, and
coagulation. An elevation of the total SOFA score of 2 points or more indicates organ dysfunction [
2
,
8
].
Sepsis is initiated by an infection [
2
]. The pathogen triggers an immune response, comprising
pro-inflammatory mechanisms to defeat the pathogen and regenerate the affected tissue, as well
as subsequent anti-inflammatory mechanisms to counteract the pro-inflammatory actions in order
to limit collateral damage in healthy tissue [
9
,
10
]. A dysregulation of this immune response, as
it appears during sepsis, leads to an over-reaction of the immune system, which can affect both
mechanisms described. Hyper-inflammation in the form of a systemic inflammatory response
syndrome (SIRS) can lead to a damage of the host’s own tissue. Immune-suppression, also known
as compensatory anti-inflammatory response syndrome (CARS), leaves the host more vulnerable
to secondary infections [
2
,
11
,
12
]. A wealth of literature is provided about sepsis and its symptoms,
diagnostics, and possible medical treatment approaches (e.g., [
10
,
11
,
13
,
14
]), to which the reader is
referred for more detailed information on these aspects of sepsis.
1.2. Zinc
Zinc is an essential trace element [
15
,
16
]. In the body it functions, for example, as a co-factor for a
high number of enzymes or as a structural element for a variety of proteins [
17
]. Zinc deficiency can
result in growth retardation, dermatitis, and hypogonadism, or symptoms such as delayed wound
healing, thymic atrophy or lymphopenia, and high incidence of infection; the latter points are due
to its particular importance for the immune system [
18
–
20
]. Consequently, zinc deficiency results
in multiple immunological changes, including what seems to be a shift toward a predominantly
innate immune response when the availability of zinc is limited [
21
]. One particularly important
effect of zinc is a modulation of the production of inflammatory cytokines [
22
]. Moreover, zinc is
crucial for the functioning of virtually all immune cells. For example, the differentiation of immature
T-cells depends on zinc, because thymulin, a hormone involved in T-cell differentiation, depends
on zinc as a co-factor [
23
,
24
]. In addition, the maturation of T-cells is influenced by their zinc
status. On the one hand a deficiency results in altered ratios of Th1- and Th2-cells, an increased
apoptosis-rate of immature T-cells, and consequently a decrease in T-cells in total [
21
,
25
–
27
]. On the
other hand, zinc supplementation has also been shown to promote regulatory T-cell development
and to suppress the maturation of Th17-cells, therefore having an inhibitory effect on Th17-mediated
autoimmune-diseases [28–30].
On the molecular level, some functions of zinc have been linked to its role as a second messenger
in immune cells. It has been shown that alterations in the intracellular free zinc-concentration function
as a “zinc signal”. Such a change in the intracellular free zinc concentration is induced by the binding
of various ligands to their respective receptors, such as lipopolysaccharide (LPS) to Toll-like receptor
4 (TLR-4), or the corresponding antigens to immunoglobulin E when it is present on the high-affinity
immunoglobulin E-receptor (Fc
ε
RI). Different kinds of immune cells vary in their expression of
receptors that utilize zinc; consequently zinc signals mediate diverse events, for example, formation
of pro-inflammatory cytokines by monocytes [
31
], presentation of major histocompatibility complex
(MHC) class II molecules at the surface of dendritic cells [
32
], formation of neutrophil extracellular
traps by neutrophil granulocytes [33], or proliferation of T-cells [34].
The essentiality of zinc for the immune system has been known since the 1960s and the
corresponding mechanistic knowledge has been expanding ever since. Its importance for the immune
system is based on various different mechanisms, each in its own way essential to ensure the
functionality of the immune system and the accurate processes of immune response, especially for
inflammatory processes. As a complete summary would exceed the scope of this article, the reader
is referred to recent review articles on the subject of zinc and immunity for more comprehensive
information [35,36] as well as to a recent review on the protective role of zinc during sepsis [37].

Nutrients 2018,10, 976 3 of 17
2. Zinc Homeostasis during Sepsis
Zinc has not only a crucial role in ensuring a proper immune response. Another observation
in the context of zinc and the immune system is an altered zinc homeostasis of the host during an
infection, which is discussed below.
2.1. Changes in Zinc Homeostasis
The host’s response to an infection or injury is referred to as an acute phase reaction (APR).
This process aims to defeat the insult, take actions against ongoing tissue damage, and re-establish
homeostasis. One of the characteristics of APR is hypozincemia. To study the time course of
hypozincemia and examine possible underlying mechanisms, Gaetke et al. injected LPS to healthy
volunteers in order to induce an inflammatory response. Subsequently, an increase in serum tumor
necrosis factor
α
(TNF-
α
) and interleukin-6 (IL-6) was observed, followed by a decrease in serum
zinc concentrations. To explain hypozincemia in their model of infection the authors suggested an
internal redistribution of zinc, mediated by cytokines [
38
]. The analysis of serum from sepsis patients
in the intensive care unit (ICU) revealed that serum zinc concentrations were reduced compared
to a healthy control group or the normal physiological range [
39
–
41
]. Probably these differences
were not caused by a zinc-deficient state due to malnutrition, but redistribution of zinc within the
patients’ bodies. Consistently, a study by Hoeger et al. showed a time-dependent decline of the
serum zinc concentrations after induction of sepsis in a porcine model [
42
]. In order to reveal the
mechanisms responsible for the observed hypozincemia, Luizzi et al. used a mouse model and
induced inflammation either by turpentine or LPS. Zrt-, Irt-like protein (ZIP)14 mRNA was the
transporter transcript that was upregulated the most. This upregulation was liver-specific and an
increase of ZIP14 on the plasma membrane of hepatocytes was shown. Further studies indicated a
role of the inflammatory cytokines IL-6 and IL-1
β
in the upregulation of ZIP14. Also, an increase in
metallothionein (MT)-1 mRNA in the liver has been observed [
43
,
44
]. This observation of enhanced
MT expression has been described before in the context of APR [45–47]. As reviewed in detail before,
MTs function as intracellular metal-binding proteins and are crucial to maintain the intracellular zinc
homeostasis. Their expression is induced by a number of metals, one of them zinc [
48
]. The increased
liver zinc concentrations accompanying hypozincemia lead to an enhanced need for zinc-binding
proteins in order to ensure the intracellular zinc homeostasis. The production of MT in the liver
seems to be regulated by cytokines as well as zinc [
49
]. Using a murine model, the already-described
decline in serum zinc concentration and an increase in liver zinc level were observed after induction
of sepsis. The analysis of the time-course of mRNA expression in the liver first showed a successive
upregulation of ZIPs 4, 6, and 10 within the first day, which then returned to near normal levels at 72 h
after induction of sepsis. The mRNA expression of ZIP14 increased at 9 h and stayed upregulated for
the time of the investigation (72 h), which is in line with the observations described previously and
supports a major role for ZIP14 in the redistribution of zinc during sepsis [
50
]. Taken together, the
APR comprises a fundamental change in liver zinc homeostasis and apparent zinc deficiency in the
serum [50,51].
The studies mentioned so far aimed, among other things, on understanding the mechanisms
responsible for the observed hypozincemia and to track the redistribution of zinc in the body.
An alternative approach to broaden the understanding of how certain processes are changed during
sepsis is gene expression analysis. The method allows insights into the impact of sepsis at the
translational level. The analysis of blood samples from pediatric septic shock patients showed a
regulation of genes that are involved in a large number of signaling pathways and gene networks,
especially those related to immunity and inflammation. Also, it has been shown that up to 12% of
the gene probes that showed a significantly decreased expression compared to the control group
are associated with the categories of “zinc, zinc finger, metal-binding and zinc-ion binding” [
52
].
These results suggest a repression of genes involved in zinc homeostasis, or depending on an intact
zinc homeostasis, as a significant feature of pediatric septic shock [
52
–
55
]. Further, the question arises
Nutrients 2018,10, 976 4 of 17
as to whether differences in gene expression in pediatric septic shock survivors and non-survivors
can be observed. Regarding zinc homeostasis, two isoforms of MT have been identified that showed
an increased expression in non-survivors compared to survivors. In addition, non-survivors had a
significantly lower serum zinc concentration compared to survivors. Considering the zinc-binding
properties of MT, Wong and colleagues interpret these results to indirectly imply that increased MT
expression in non survivors might affect zinc homeostasis and thereby serum zinc concentration [
53
].
Taken together, different approaches imply a contribution of zinc and its altered homeostasis to
the pathobiology of sepsis.
2.2. Possible Reasons for the Redistribution of Zinc
With respect to the considerable differences of serum zinc concentrations between sepsis patients
and the corresponding control groups, as well as the finely tuned alterations of zinc homeostasis, it
can be assumed that these are part of a directed process that aims to benefit the host in defeating the
pathogen. This process includes a decrease in serum zinc concentration as well as an increase in liver
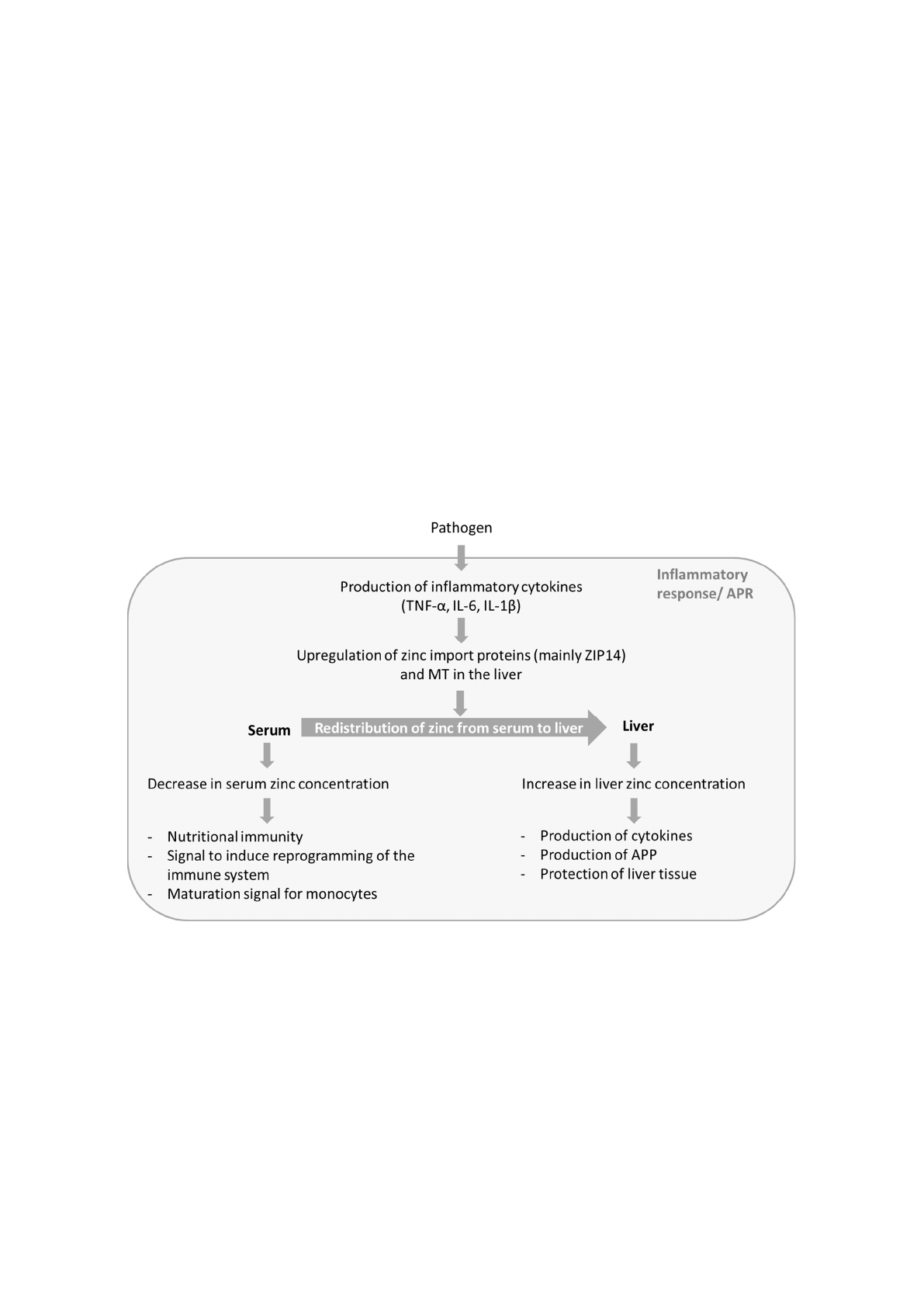
zinc concentration, whereas both aspects seem to benefit the host’s defense against pathogens. Figure 1
gives a brief overview of the processes causing the alterations in zinc concentrations, as well as the
possible beneficial effects.
Nutrients 2018, 10, x FOR PEER REVIEW 4 of 17
increased expression in non-survivors compared to survivors. In addition, non-survivors had a
significantly lower serum zinc concentration compared to survivors. Considering the zinc-binding
properties of MT, Wong and colleagues interpret these results to indirectly imply that increased MT
expression in non survivors might affect zinc homeostasis and thereby serum zinc concentration [53].
Taken together, different approaches imply a contribution of zinc and its altered homeostasis to
the pathobiology of sepsis.
2.2. Possible Reasons for the Redistribution of Zinc
With respect to the considerable differences of serum zinc concentrations between sepsis
patients and the corresponding control groups, as well as the finely tuned alterations of zinc
homeostasis, it can be assumed that these are part of a directed process that aims to benefit the host
in defeating the pathogen. This process includes a decrease in serum zinc concentration as well as an
increase in liver zinc concentration, whereas both aspects seem to benefit the host’s defense against
pathogens. Figure 1 gives a brief overview of the processes causing the alterations in zinc
concentrations, as well as the possible beneficial effects.
Figure 1. Possible functions of zinc in sepsis. During the APR of sepsis zinc is redistributed from
serum to liver. This process results in decreased serum zinc concentration and increased liver zinc.
The altered zinc concentrations seem to serve different functions and to be a part of the host’s
defense against pathogens. APR: acute phase reaction; IL: interleukin; TNF: tumor necrosis factor;
MT: metallothionein; APP: acute phase proteins.
Research on zinc homeostasis in the context of sepsis delivered a variety of explanations for the
beneficial effects of a redistribution of zinc. The respective studies are discussed below.
One of the main effects of the redistribution of zinc is an accumulation of zinc in the liver.
Hence, it seems as is if a higher liver zinc level might benefit the host during infection. Among other
things, the APR is not only characterized by the previously mentioned redistribution of zinc, but also
by production of acute phase proteins (APP) and the release of cytokines [56,57]. Zinc serves as an
important structural element for many proteins and is required by enzymes involved in
transcription and translation. Therefore, the higher synthesis rate of APP in the liver could cause an
increased requirement for zinc during APR [56–58]. With respect to cytokine production a knockout
(k.o.) of ZIP14 in mice, a transporter important for the regulation of zinc homeostasis in hepatocytes,
showed lower mRNA expression of TNF-α, IL-6, IL-1β, and IL-10 in the liver compared to wild-type
Figure 1.
Possible functions of zinc in sepsis. During the APR of sepsis zinc is redistributed from serum
to liver. This process results in decreased serum zinc concentration and increased liver zinc. The altered
zinc concentrations seem to serve different functions and to be a part of the host’s defense against
pathogens. APR: acute phase reaction; IL: interleukin; TNF: tumor necrosis factor; MT: metallothionein;
APP: acute phase proteins.
Research on zinc homeostasis in the context of sepsis delivered a variety of explanations for the
beneficial effects of a redistribution of zinc. The respective studies are discussed below.
One of the main effects of the redistribution of zinc is an accumulation of zinc in the liver.
Hence, it seems
as is if a higher liver zinc level might benefit the host during infection. Among other
things, the APR is not only characterized by the previously mentioned redistribution of zinc, but also
by production of acute phase proteins (APP) and the release of cytokines [
56
,
57
]. Zinc serves as an

Nutrients 2018,10, 976 5 of 17
important structural element for many proteins and is required by enzymes involved in transcription
and translation. Therefore, the higher synthesis rate of APP in the liver could cause an increased
requirement for zinc during APR [
56
–
58
]. With respect to cytokine production a knockout (k.o.) of
ZIP14 in mice, a transporter important for the regulation of zinc homeostasis in hepatocytes, showed
lower mRNA expression of TNF-
α
, IL-6, IL-1
β
, and IL-10 in the liver compared to wild-type (w.t.)
mice after induction of sepsis in a murine model. Simultaneously, plasma levels of TNF-
α
, IL-6,
and IL-10 were significantly higher in k.o. than in w.t. mice. The results indicate a disadvantage of
the ZIP14 k.o. mice during sepsis based on increased markers of inflammation and an influence of
zinc, transported by ZIP14, on the production of cytokines during APR. However this observation is
surprising, because a decrease in mRNA expression would be expected to result in lower cytokine
levels. Possible explanations would be elevated cytokine expression elsewhere in the body, or an
impact on mechanisms mediating the expression of antagonists of the pro-inflammatory cytokines [
50
].
Other studies suggest that the altered zinc supply during endotoxemia has a major influence
on energy production in the liver. Injection of LPS caused an increase in hepatic zinc and MT in w.t.
mice, whereas zinc levels stayed unchanged in MT k.o. mice. At the same time the liver glucose of the
former stayed unchanged while the levels in MT k.o. mice decreased significantly. These results imply
a lack of hepatic gluconeogenesis in the MT KO mice and a role for MT, and most likely also for zinc,
in maintaining glycaemia after induction of an infection [59].
A protective role of zinc for the liver has also been suggested. Using murine models it was shown
that after injection of endotoxin, zinc-deficient nutrition resulted in enhanced lipid peroxidation in
the liver compared to the zinc adequate group [
60
]. Another study showed that zinc pre-treatment of
mice resulted in an increased intracellular availability of zinc in liver cells and was accompanied
by decreased accumulation of superoxide and necrotic cell death in the liver after injection of
LPS [
61
]. Both experimental observations support a protective role of zinc in the liver during infection.
Interestingly, after injection of endotoxin, Sakagouchi et al. saw an increase in MT only in the zinc
adequate group, but not in zinc-deficient animals, and therefore suggested a relation between zinc
concentration, endotoxin-induced MT and lipid peroxidation [
60
]. In contrast, Zhou et al. observed the
protective effects of zinc pre-treatment on liver cells in w.t. mice as well as MT k.o. mice, leading them
to propose an effect of zinc independent of MT [
61
]. These results are not necessarily contradictory,
but could imply that the protective effect of zinc on the liver could work in more than one way.
Further studies on this topic would be useful, since organ dysfunction is one of the hallmarks of
sepsis and a better understanding of its mechanisms is the basis of a possible prevention. In summary,
studies show multiple and diverse functions of zinc in the liver during the onset of sepsis, suggesting
a physiological basis for the accumulation of zinc.
The redistribution of zinc and accumulation in the liver is accompanied by a decrease in serum
zinc concentration. With regard to the host’s defense against pathogens, this effect might have some
benefits as well. One protective mechanism of the host is referred to as nutritional immunity. Pathogens,
just like all living organisms, require transition metals for their survival. The host’s strategy is to restrict
the pathogens’ access to essential transition metals, for example by lowering their concentrations in
the serum or secretion of metal ion binding proteins. This process is not limited to zinc but has been
described for other micronutrients, such as iron or manganese [62].
A decrease in the serum zinc concentration has also been shown to influence the respective
number and maturation of immune cells. Therefore, the alteration of serum zinc during the APR might
function as a signal. Using a murine model of zinc deficiency, a downregulation of lymphopoiesis
and upregulation of myelopoiesis was found [
19
,
21
,
27
,
63
]. In line with this observation, a decrease
of intracellular zinc occurred as a result of homeostatic changes during monocytic differentiation of
HL60 cells. Moreover, experiments simulating zinc deficiency showed that lower zinc levels promoted
the development of HL-60 cells along the myeloid lineage into functionally mature macrophages [
64
].
The immune cells that benefit from a decline of serum zinc are part of the innate immune system.
They represent the first line of host defense and provide a faster response than the cells of the adaptive
Loading more pages...