
J Appl Ecol. 2020;57:1581–1592.
|
1581wileyonlinelibrary.com/journal/jpe
Received: 2 March 2020
|
Accepted: 8 April 2020
DOI: 10.1111/1365-2664.13661
RESEARCH ARTICLE
Plant traits, biotopes and urbanization dynamics explain the
survival of endangered urban plant populations
Greg Planchuelo1 | Ingo Kowarik1,2 | Moritz von der Lippe1,2
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium,
provided the original work is properly cited.
© 2020 The Authors. Journal of Applied Ecology published by John Wiley & Sons Ltd on behalf of British Ecological Society
1Department of Ecology, Ecosystem
Science/Plant Ecology, Technische
Universität Berlin, Berlin, Germany
2Berlin-Brandenburg Institute of Advanced
Biodiversity Research, Berlin, Germany
Correspondence
Greg Planchuelo
Email: greg.planchue[email protected]
Funding information
DAAD / Technische Universität Berlin; Hans
Böckler Stiftung, Grant/Award Number:
398226
Handling Editor: Peter Manning
Abstract
1. With accelerating urbanization, the urban contribution to biodiversity conservation
becomes increasingly important. Previous research shows that cities can host
many endangered plant species. However, fundamental questions for urban na-
ture conservation remain open: to what extent and where can endangered plant
species persist in the long term and which mechanisms underlie population
survival?
2. We evaluate the survival of 858 precisely monitored populations of 179 endan-
gered plant species in Berlin, Germany, by assessing population survival through-
out different urban ecosystems over a period of 7.6 years on average. By linking
population survival to various landscape variables and plant traits, we unravel the
underlying drivers.
3. More than one–third of populations went extinct during the observation period.
Population survival was inversely correlated to the increase in impervious sur-
faces in the vicinity following the first 11 years after the fall of the Berlin wall.
Additionally, populations in semi-natural habitats like forests and bogs were sur-
prisingly more prone to local extinction than populations in anthropogenic habi-
tats. Survival was highest for competitive species with a preference for drier soils
(Ellenberg indicator for soil humidity).
4. Synthesis and applications. Considerable levels of local population extinction dem-
onstrate that the presence of endangered plants cannot be directly linked with
their long-term survival in cities. However, the survival of remaining populations
indicates opportunities for urban biodiversity conservation both within and out-
side conservation areas. The elucidated links between population survival, ur-
banization dynamics, biotope class and species traits support urban conservation
strategies that reduce the proportion of impervious surface, prioritize conserva-
tion management in forests and grasslands and explore the opportunities of green
spaces and built-up areas.
KEYWORDS
anthropogenic biotopes, biodiversity conservation, endangered plant species, plant traits,
population monitoring, population persistence, urban ecosystems, urbanization changes

1582
|
Journal of Applied Ecology
PLANCHUELO Et AL.
1 | INTRODUCTION
As urbanization accelerates globally, the question of the urban
contribution to biodiversity conservation is becoming increasingly
important (Kowarik & von der Lippe, 2018; McKinney, 2002; Nilon
et al., 2017; Parris et al., 2018). The number of urban conservation
studies has risen sharply in recent years (Shwartz, Turbé, Julliard,
Simon, & Prévot, 2014) and indicate considerable opportunities for
biodiversity conservation. Indeed, cities can be very rich in native
species (Aronson et al., 2014), including a considerable richness of
endangered plant species as reported from cities in Africa (Rebelo,
Holmes, Dorse, & Wood, 2011), Asia (Wang et al., 2007), Australia
(Ives et al., 2016), Europe (Kowarik & von der Lippe, 2018) and North
America (Lawson, Lamar, & Schwartz, 2008). From these insights, a
narrative regarding cities' contribution to biodiversity conservation
has developed, and with this several strategies to promote urban
biodiversity have been suggested. Classical approaches such as the
designation of protected areas are increasingly complemented by
integrative approaches arguing for a biodiversity-friendly manage-
ment of green spaces (Aronson et al., 2017; Chollet, Brabant, Tessier,
& Jung, 2018), a reconciliation of urban land use and biodiversity
conservation (Elmqvist et al., 2013), the integration of biodiversity in
urban design (Garrard, Williams, Mata, Thomas, & Bekessy, 2018) or
the promotion of urban wilderness (Hwang, Yue, Ling, & Tan, 2019;
McKinney, Kowarik, & Kendal, 2018).
However, strategies to promote urban biodiversity in general
may not specifically support the protection of endangered plant
species. While target species conservation is a major motive for
urban biodiversity conservation (Dearborn & Kark, 2010), the meta-
analysis by Shwartz et al. (2014) revealed that empirical evidence
for the effectiveness of urban conservation strategies is lacking. It
therefore remains unclear which criteria should guide the allocation
of scarce resources for urban nature conservation or whether the
contribution of cities to the conservation of biodiversity is generally
overestimated (Shwartz et al., 2014). An assessment of the long-term
survival of endangered plant species in cities is currently hampered
by knowledge deficits in three key areas.
1.1 | Population persistence
The occurrence of species of conservation concern in cities does not
necessarily indicate their effective protection (Shwartz et al., 2014).
Environmental constraints due to urban land use, fragmentation or
novel environmental stressors represent significant limitations on
plant dispersal and recruitment (Kowarik & von der Lippe, 2018;
McDonnell & Hahs, 2015; Williams et al., 2009), while habitat loss
and change can be responsible of local plant extinctions (Duncan
et al., 2011; Knapp, Kühn, Stolle, & Klotz, 2010). Thus, population
persistence is a key issue for sustainable target species conserva-
tion in cities, as there is evidence for decreasing population sizes
in urban plants over time (Chocholoušková & Pyšek, 2003; Knapp
et al., 2010), and remaining populations of a species can thus mask
future extinction debts (Hahs & McDonnell, 2014). However, most
urban biodiversity studies report as snapshot studies the occurrence
of species at a certain point in time, but do not allow conclusions
to be drawn about the persistence of urban populations (but see
Lawson et al., 2008; Schwartz, Smith, & Steel, 2013).
1.2 | Type of habitat
While the occurrence of target species of conservation concern
across different habitats in cities has often been described, the
relative importance of different urban habitats for such species is
not fully understood (Shwartz et al., 2014). Species of conserva-
tion concern have been reported from very different habitat types
within cities (Planchuelo, von der Lippe, & Kowarik, 2019), for ex-
ample, forests (Godefroid & Koedam, 2003), brownfields (Bonthoux,
Brun, Di Pietro, Greulich, & Bouché-Pillon, 2014), parks (Cornelis &
Hermy, 2004) or cemeteries (Kowarik, Buchholz, von der Lippe, &
Seitz, 2016; Löki, Deák, Lukács, & Molnár, 2019). However, an im-
portant bias is that populations of endangered species have not been
studied equally across all urban habitat types. Rather, most urban
conservation studies address larger habitats (roughly >2 ha), with
natural remnants and large green spaces being overrepresented
(Shwartz et al., 2014). Moreover, species inventories of urban nature
reserves are likely better known than those of unprotected land use
types.
1.3 | Environmental and urbanization predictors
Which species occur in urban habitats not only depends on local
habitat features such as patch size but also on features of the sur-
rounding urban matrix as indicated by urbanization parameters (e.g.
the proportion of impervious surface, human population density)
or the vicinity to (semi-)natural ecosystems. Previous urban studies
have revealed the relative importance of many potential predictors
of species richness (e.g. Anderson & Minor, 2019; Beninde, Veith,
& Hochkirch, 2015; Westermann, von der Lippe, & Kowarik, 2011;
Williams, Morgan, McCarthy, & McDonnell, 2006). However, results
based on total species richness cannot necessarily be generalized
to endangered plant species, as shown for urban grassland re-
serves in Prague (Jarošík, Konvička, Pyšek, Kadlec, & Beneš, 2011).
Specific analyses for endangered plant species are rare and yielded
ambiguous results: studies at larger national scales revealed posi-
tive relationships between urbanization parameters and species of
conservation concern (Kühn, Brandl, & Klotz, 2004; Lenzen, Lane,
Widmer-Cooper, & Williams, 2009; Shwartz, Muratet, Simon, &
Julliard, 2013), while studies at smaller city scales yielded contrast-
ing results (Schmidt, Poppendieck, & Jensen, 2014). However, many
studies rely on heterogeneous urban landscapes (e.g. grids) without
specifying the habitat scale, that is, the specific locations of endan-
gered plant populations. Moreover, dynamics of urban environments
are an important features of cities (Ramalho & Hobbs, 2012). Change

|
1583
Journal of Applied Ecology
PLANCHUELO Et AL.
rates of urbanization measures (e.g. impervious surface, population
size) can determine urban biodiversity patterns (Fischer, Rodorff,
von der Lippe, & Kowarik, 2016). Such measures are rarely incorpo-
rated into analyses of the long-term survival of endangered species,
although dynamic changes of urban environments can potentially
threaten population survival, for example, by loss of open spaces, or
indirect urbanization effects.
1.4 | Species traits
Besides characteristics of a population's environment, species iden-
tity or more generally, species traits, are held responsible for the vul-
nerability to local extinction in cities (Duncan et al., 2011). Although
several studies have indicated shifts in specific plant traits due to
urbanization, there was only a marginal impact of species traits on
plant extinctions in another study (Duncan et al., 2011). However,
higher frequency in urban habitats has been linked to competitive
traits (e.g. plant height; Fischer, von der Lippe, & Kowarik, 2013) or
strategies (Chocholoušková & Pyšek, 2003), and to preference for
nutrient-rich sites (Knapp et al., 2009), while lower frequencies in
cities are reported for species with high water requirements (Knapp
et al., 2010).
We aimed to elucidate the persistence of endangered plant spe-
cies in cities—and underlying mechanisms—by evaluating a unique
dataset from Berlin. These data contained long-term monitoring
results of a large number of precisely mapped plant populations
(n = 858) belonging to 179 endangered plant species. These plant
species have been identified as priority species of conservation
concern in Berlin's Flora Protection Program and the extant popu-
lations of these species have been subjected to a repeated monitor-
ing (Meißner & Seitz, 2010). Therefore, information was available on
which populations have survived or have gone locally extinct at the
respective sites since the first survey.
To unravel mechanisms that might underpin population survival,
we assigned each of the originally mapped populations (a) to a spe-
cific biotope type and (b) characterized the site occupied by each
population by a range of urban landscape variables whose funda-
mental relevance has been shown previously and that were related
to habitat features, the urbanization of the surroundings and the
vicinity of (near)-natural ecosystems. Additionally, we linked popu-
lation survival to plant traits that have been shown to be relevant to
plant performance in previous urban studies.
We expected population survival rates to decrease with increas-
ing urbanization or rapid changes in the areas surrounding each pop-
ulation and to differ significantly among biotope types. We assumed
a positive correlation between population survival and an increasing
proximity to natural remnants or large forest and grassland areas,
which might act as propagule sources. Furthermore, we expected
a positive relationship between plant traits related to competitive
ability and population survival.
In detail, we addressed the following research questions: How is
the survival rate of populations of endangered plant species in Berlin
related to (a) features of the respective habitats (e.g. type of biotope,
patch size, legal conservation status); (b) the degree and dynamics
of urbanization of the surrounding urban matrix (e.g. current share
and change in impervious surface area, human population density);
(c) to the vicinity of (near)natural ecosystems (e.g. distance to natural
remnants, proportion of forests in buffer around the sites) and (d)
plant functional traits of the respective endangered plant species
(e.g. Ellenberg indicators, CSR strategy).
2 | MATERIALS AND METHODS
2.1 | Study area
This study was carried out in Berlin, the largest city of Germany, with
3.6 million inhabitants in 2017. Berlin is a typical large European
city, including a wide range of ecosystems ranging from remnants
of semi-natural and agrarian landscapes or urban greenspaces with
different land use histories to novel ecosystems in vacant urban-
industrial land. Green and blue spaces make up 41% of the total
area of Berlin, which is 891 km2; the remaining 59% is covered by
built-up and traffic areas (SenStadtUm, 2016). Red lists of endan-
gered species were established in the 1980s and updated several
times since. From today's perspective, we thus know that 17% of
Berlin's flora has gone extinct since the mid-19th century, and 29%
is currently being endangered (Seitz, Ristow, Meißner, Machatzi, &
Sukopp, 2018).
2.2 | Population data
We used an extensive monitoring dataset from Berlin's Flora
Protection Program (Berliner Florenschutzkonzept) on the precise
geographical location of 858 populations of 179 endangered plant
species in Berlin (see Appendix S1). From these, 72.5% of the spe-
cies are herbaceous, 20.6% geophytes, 3.5% shrubs, 1.9% hydro-
phytes and 1.5% trees (Table 1). The 10 most populous plant families
are, in decreasing order, Rosaceae, Cyperaceae, Caryophyllaceae,
Asteraceae, Poaceae, Orchidaceae, Ranunculaceae, Iridaceae and
Fabaceae (Table 1). The monitored species have the highest con-
servation priority because they are threatened locally and/or at
regional, national or global scales (Meißner & Seitz, 2010). A popula-
tion was discerned from another when individuals where separated
at least 30 m or when populations were clearly separated by roads
or paved paths. We used the dataset in previous studies (Planchuelo,
Kowarik, & von der Lippe, 2020a; Planchuelo et al., 2019) but here
include for the first time information on population survival. The first
inventory of populations was compiled from several expert surveys
after 1990. On the basis of the georeferenced locations of this initial
dataset, a re-mapping project took place between 2009 and 2014,
aiming to relocate all previously mapped populations. The average
period of time between the first and second mapping of a population
was 7.6 (±0.2) years. This allowed us to differentiate persistent from
1584
|
Journal of Applied Ecology
PLANCHUELO Et AL.
locally extinct populations of those recorded in the initial dataset.
To avoid methodological bias, we excluded all populations from the
initial compilation with imprecise information on the spatial location,
and populations in open waters, due to difficulties in relocating and
resurveying them.
2.3 | Habitat features
To evaluate the role of different biotope types for population sur-
vival, we intersected the location of each mapped population with
the respective biotope class from the Berlin biotope mapping
(SenStadtUm, 2014). Terrestrial populations of endangered plant
species were present in eight major biotope classes (Figure 1): for-
ests, grasslands, ruderal sites, built-up areas, bogs and marshes,
groves and hedges, green spaces and agricultural fields. We calcu-
lated the area of the respective biotope patches to elucidate the role
of patch size for population survival.
To test for the role of the legal protection status of an area on
population survival, we attributed the site of each population to
one of the following categories: nature reserve (Naturschutzgebiet),
Natura 2000 site, protected landscape (Landschaftsschutzgebiet)
and unprotected sites. The first three categories represent legally
protected areas, with decreasingly strict protection status and man-
agement measures. We also assessed whether the populations were
located in former East Berlin or West Berlin to evaluate if the differ-
ent patterns of urban development in both parts of the city had an
effect on population survival.
2.4 | Landscape variables
We determined several characteristics of the urban matrix surround-
ing the populations of endangered plants and used them to reveal the
effects of urbanization on population survival (see Appendix S2 for
parameters and sources). We calculated data on the proportion of
impervious surface, the mean floor space index (a measure of urban
TABLE 1 Number of populations and species belonging to
different life forms and to the 10 most populous plant families of
the endangered plants used in our study
Number of
populations
Number of
species
Life form
Herbaceous perennial 566 66.0% 101 56.4%
Geophyte 177 20.6% 30 16.8%
Herbaceous annual 56 6.5% 23 12.8%
Shrub 30 3.5% 15 8.4%
Hydrophyte 16 1.9% 95.0%
Tree 13 1.5% 10.6%
Plant family
Rosaceae 96 11.2% 13 7.3%
Cyperaceae 89 10.4% 15 8.4%
Caryophyllaceae 84 9.8% 11 6.1%
Asteraceae 83 9.7% 21 11.7%
Poaceae 73 8.5% 13 7.3%
Orchidaceae 47 5.5% 84.5%
Ranunculaceae 44 5.1% 10 5.6%
Iridaceae 35 4.1% 10.6%
Fabaceae 28 3.3% 63.4%
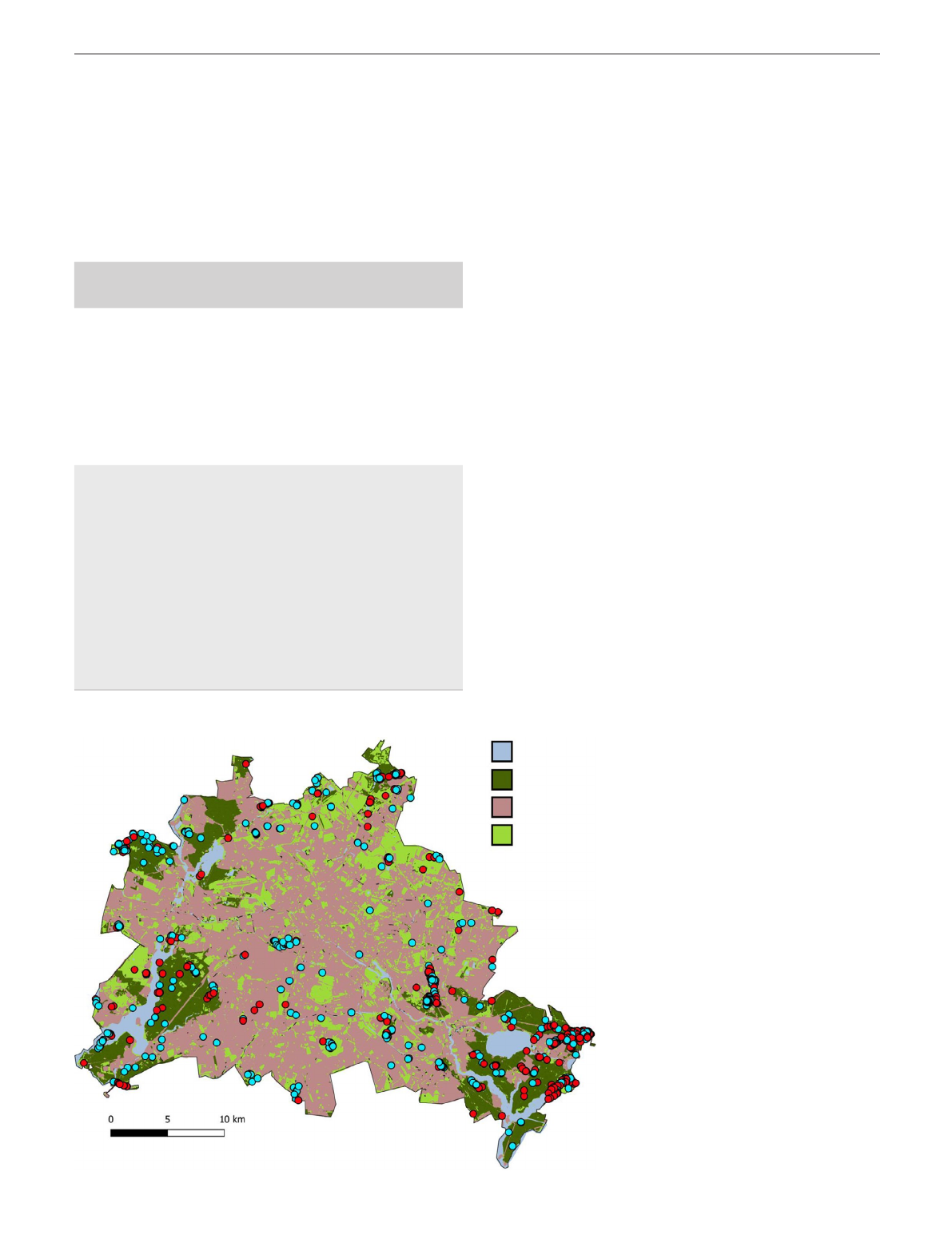
FIGURE 1 Location of populations
(n = 858) of endangered plant species in
Berlin that survived (blue dots) or did not
survive (red dots) during the monitoring
period. The category ‘open land’ includes
biotope classes that were further
differentiated in the analyses
Water bodies
Forests
Built up areas
Open land

|
1585
Journal of Applied Ecology
PLANCHUELO Et AL.
density, i.e. the ratio of a buildings total floor area and its lot area),
human population density and the length of the road network in a
500 m radius buffer surrounding each population of endangered plant
species. To account for effects of spatial isolation from (near)natural
ecosystems, we calculated the proportion of forests and grasslands
for a 500 m radius buffer around each population, based on informa-
tion from the Berlin biotope map (SenStadtUm, 2014).
To account for urbanization dynamics throughout the monitor-
ing period, we calculated the changes in the proportion of imper-
vious surface area throughout two periods of time (i.e. 1990–2001
and 2005–2016) for a 500 m radius buffer around each population.
These periods reflect different phases of urbanization: the first de-
cade after the fall of the Berlin Wall in 1989, and a period of less dy-
namic but progressive urban development. Both periods are covered
by citywide measurements of the impervious surface area using the
same method. As a measure of socioeconomic change, we compiled
mean human population constancy (i.e. the percentage of residents
living at least 5 years in the same address) in the same buffer from
the Berlin map of land use (SenStadtUm, 2016).
To calculate the average values of each parameter, we spatially inter-
sected the corresponding map with each individual buffer in QGIS (2019)
and then used the plugin ‘Dissolve with Stats’ to obtain the average val-
ues for each buffer. The distance of each population to the nearest road
and to the nearest natural remnant was calculated in QGIS (2019) with
the ‘NN Join’ plugin. We addressed those biotopes as natural remnants
if they corresponded to historical natural ecosystems, being only slightly
affected by human impacts, following the classification by Kowarik and
von der Lippe (2018). Such remnants include mires, wetlands, semi-nat-
ural forests and some semi-natural grasslands. Spatial information on
natural remnants was derived from Planchuelo et al. (2019). Appendix
S3 summarizes the methodological approach of the study.
2.5 | Plant traits
Plant traits, CSR strategy types (Grime, 1977) and Ellenberg indica-
tors (Ellenberg, Weber, Düll, Wirth, & Werner, 1991) have been suc-
cessfully linked to urban biodiversity patterns in previous studies
(Williams, Hahs, & Vesk, 2015). We thus included these parameters
as predictors of population survival in our analyses (Appendix S4).
Information on plant morphological traits (SLA, plant height) and the
CSR strategy of the species was gathered from the BIOFLOR database
(Klotz, Kühn, Durka, & Briemle, 2002). For information on the realized
niche of a species, we used indicator values for the soil humidity, soil
nitrogen and soil acidity requirements, and for light and temperature
requirements of each species from Ellenberg et al. (1991).
2.6 | Statistical analyses
To relate population survival to habitat characteristics and
landscape variables (Appendix S2), we used a GLMM (R func-
tion glmer; Bates, 2010). We coded population survival as a
binomial response and corrected for spatial and phylogenetic
dependence by including the spatial coordinates of their loca-
tions (longitude and latitude) and plant genus as random fac-
tors. Plant genus has been included as a random factor in GLMM
models to successfully account for phylogenetic dependencies
(e.g. Pandit, White, & Pocock, 2014). Because of the varying pe-
riod between the first and the second monitoring of the popu-
lations, this time span in years was also included as a random
effect in our model. All habitat and landscape variables were
tested for intercorrelation and as no correlations higher than
|r| < 0.7 appeared, all of them were included as fixed effects in
a full model (Dorman et al., 2013). A minimal adequate model
was chosen by stepwise backward selection of the predictors
based on minimal AIC. Random variable selection was done
separately with the entire set of variables prior to backward
selection of the fixed effects. We evaluated the prediction ac-
curacy of the model by calculating the area under the receiver
operating curve (AUC) with the package pROC in R (Robin et al.,
2011).
To estimate the effect of plant traits on population survival
and to account for possible nonlinear effects, we calculated the
relative importance of each variable through a conditional ran-
dom forest and successively performed a conditional inference
tree analysis. For both, we used the same response as in the
GLMM (binomial population survival) and the set of predictors
related to the morphological plant traits of the species, their
CSR strategy and their realized niche as expressed through
Ellenberg indicator values (Ellenberg et al., 1991; Appendix S4).
A variable selection through a conditional random forest is a
machine learning technique that is able to estimate the relative
importance of highly correlated predictors (Strobl, Boulesteix,
Kneib, Augustin, & Zeileis, 2008). A conditional inference tree
is a non-parametric type of decision tree where the dataset is
recursively split into dichotomous subsets, which are discrim-
inated by the most significant predictor (Hothorn, Hornik, &
Zeileis, 2006). We chose these models because of their ability
to display interactions between different traits and their flexible
handling of categorical data.
All analyses were performed with the statistical and program-
ming software R version 3.5.2 (R Core Team, 2018).
3 | RESULTS
3.1 | Survival rate
Almost two-thirds of the populations of endangered plant species
(64%) mapped in the first monitoring were confirmed in the second
monitoring (551 populations). Thus, more than one-third of all popu-
lations became locally extinct during the period between the two
monitoring dates, which averaged 7.6 years. During that time, from
a total of 179 endangered species, 49 (27%) went extinct for the
study sites.
Loading more pages...