
full paper
© 2017 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
wileyonlinelibrary.com
1606265 (1 of 9)
and chloroplasts. In addition to greater
control over compartmentalization, the
extra layering of membranes gives the
organisms increased area for membrane
proteins. The inner membranes of mito-
chondria, for example, are folded into
cristae to greatly increase the surface area
available for the electron transport chain
proteins, packing high concentrations into
a small volume.[1] Similarly, chloroplasts
employ the stacking of interconnected
thylakoid membranes to greatly enhance
the concentrations of photosynthetic pro-
tein complexes required to efficiently
harness solar energy.[2] These structures
and many others show how the lamellar
stacking of membranes and their proteins
hold substantial technological potential
for biomimicry, where the stacking of
membranes may allow the design of novel
protein arrangements with possible appli-
cations in catalysis, photonics, sensing,
and the 3D crystallization of membrane
proteins.[3,4]
Supported lipid bilayers (SLBs) have been widely advocated
for biotechnological applications and frequently used as model
cell membranes in fundamental studies. SLBs provide a pow-
erful means to study the function of membrane proteins and
protein–protein interaction in membranes.[5–7] The planar
nature of SLBs allows investigations by an array of powerful
surface analytical techniques such as atomic force microscopy
(AFM), quartz crystal microbalance, surface plasmon resonance,
and total internal reflection fluorescence microscopy. The poten-
tial of SLBs to mimic complex multilayer membrane assemblies
has been reported,[8] yet the development of consistent meth-
odologies has been limited, and only few model systems are
described that investigate protein behavior in multiple mem-
branes. For instance, multilamellar membranes are routinely
used in studies of the structure of lipid membranes and mem-
brane-associated polypeptides by X-ray or neutron diffraction, or
by solid-state nuclear magnetic resonance spectroscopy.[9] Meth-
odologies used to form these multilayered membranes require
the drying of membranes in organic solvents, which is suitable
for lipid and peptide studies; however, during drying, mem-
brane proteins are prone to lose their biological activity.[10]
We have previously reported on a simple layer-by-layer
(LBL) methodology to form lipid multilayers via vesicle rup-
ture onto existing SLBs using poly-l-lysine (PLL) as an elec-
trostatic polymer “glue.”[11] Not only does this technique allow
Multilayered Lipid Membrane Stacks for Biocatalysis Using
Membrane Enzymes
George R. Heath, Mengqiu Li, Honling Rong, Valentin Radu, Stefan Frielingsdorf,
Oliver Lenz, Julea N. Butt, and Lars J. C. Jeuken*
Multilayered or stacked lipid membranes are a common principle in biology and
have various functional advantages compared to single-lipid membranes, such
as their ability to spatially organize processes, compartmentalize mole cules, and
greatly increase surface area and hence membrane protein concentration. Here,
a supramolecular assembly of a multilayered lipid membrane system is reported
in which poly-l-lysine electrostatically links negatively charged lipid membranes.
When suitable membrane enzymes are incorporated, either an ubiquinol oxi-
dase (cytochrome bo3 from Escherichia coli) or an oxygen tolerant hydrogenase
(the membrane-bound hydrogenase from Ralstonia eutropha), cyclic voltam-
metry (CV) reveals a linear increase in bio catalytic activity with each additional
membrane layer. Electron transfer between the enzymes and the electrode is
mediated by the quinone pool that is present in the lipid phase. Using atomic
force microscopy, CV, and fluorescence micro scopy it is deduced that quinones
are able to diffuse between the stacked lipid membrane layers via defect sites
where the lipid membranes are inter connected. This assembly is akin to that
of interconnected thylakoid membranes or the folded lamella of mitochondria
and has significant potential for mimicry in biotechnology applications such as
energy production or biosensing.
Dr. G. R. Heath, Dr. M. Li, H. Rong, Dr. V. Radu,
Dr. L. J. C. Jeuken
School of Biomedical Sciences
University of Leeds
Leeds LS2 9JT, UK
E-mail: [email protected]
Dr. S. Frielingsdorf, Dr. O. Lenz
Institut für Chemie, Sekretariat PC14
Technische Universität Berlin
Straße des 17. Juni 135, 10623 Berlin, Germany
Prof. J. N. Butt
Centre for Molecular and Structural Biochemistry
School of Biological Sciences and School of Chemistry
University of East Anglia
Norwich NR4 7TJ, UK
DOI: 10.1002/adfm.201606265
1. Introduction
Double or multilayered membranes, along with their asso-
ciated membrane proteins, are an integral part of the energy
producing pathways of eukaryotic cells, gram-negative bacteria,
The copyright line of this paper was changed 5 April 2017 after initial
publication.
This is an open access article under the terms of the Creative Commons
Attribution License, which permits use, distribution and reproduction in
any medium, provided the original work is properly cited.
www.afm-journal.de
Adv. Funct. Mater. 2017, 27, 1606265
www.advancedsciencenews.com
full paper
© 2017 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim wileyonlinelibrary.com (2 of 9) 1606265
incorporation of membrane proteins, but it can also create
membrane stacks with different membrane proteins in each
layer. Alternating membrane protein stacks are found in, for
example, the electrocyte cells of electric eels in which the trans-
membrane proteins are asymmetrically distributed across two
primary membranes that, when added in series, can generate
potentials of ≈600 V.[12,13]
Here, we engineered a lipid multilayer matrix in which
native electron mediators can freely diffuse for bio-electro-
catalytic applications. Mimicking the function of cristae and
thylakoid stacks in mitochondria and chloroplasts, respec-
tively, the assembly increases the concentration of membrane
enzymes per electrode surface area in a stepwise manner. We
created multilayers of membrane enzymes in a native-like
lipid environment using the LBL assembly of bacterial mem-
brane extracts at gold electrodes. Assemblies with two different
membrane proteins are demonstrated: a membrane-bound
hydrogenase (MBH) from Ralstonia eutropha, which oxidizes
H2, and a ubiquinol oxidase, cytochrome bo3, from Escherichia
coli that reduces oxygen. With each deposition of a membrane
layer, we see the catalytic activity increasing as the total amount
of enzyme on the surface increases. This biomimetic system
demonstrates how the stacking of membranes can proportion-
ally increase the concentration of active membrane proteins at
surfaces.
2. Results and Discussion
2.1. Interconnected Lipid Multilayers
Initial structural characterization of the membrane stacks was
performed on glass and mica using fluorescence microscopy
and AFM, respectively. Quinone diffusion between the mem-
brane layers and biocatalytic activity of membrane enzymes
incorporated into the stack was then characterized using
electrochemistry, for which the stacks were formed on gold
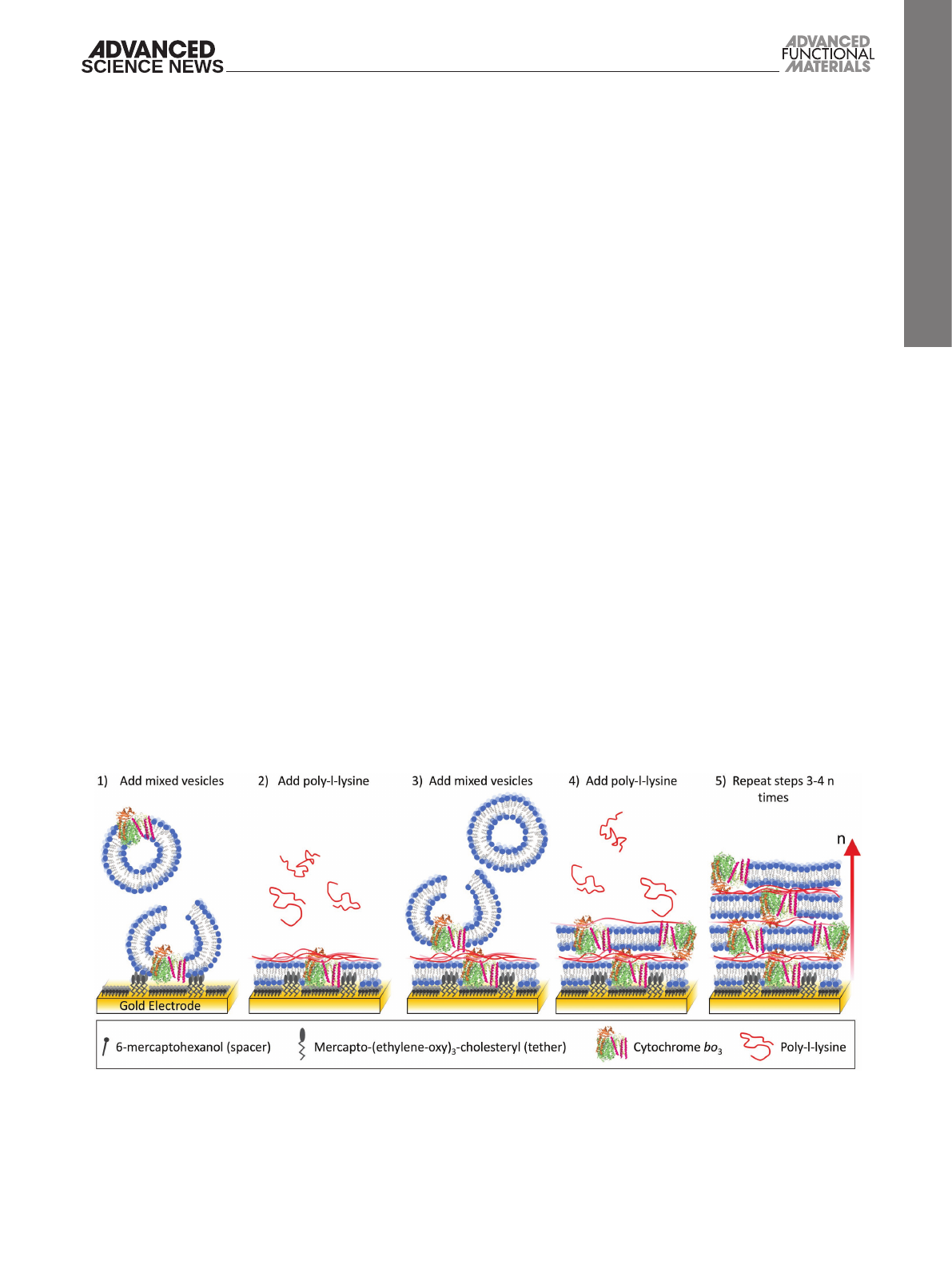
surfaces. The formation of membrane stacks is schematically
shown in Figure 1.
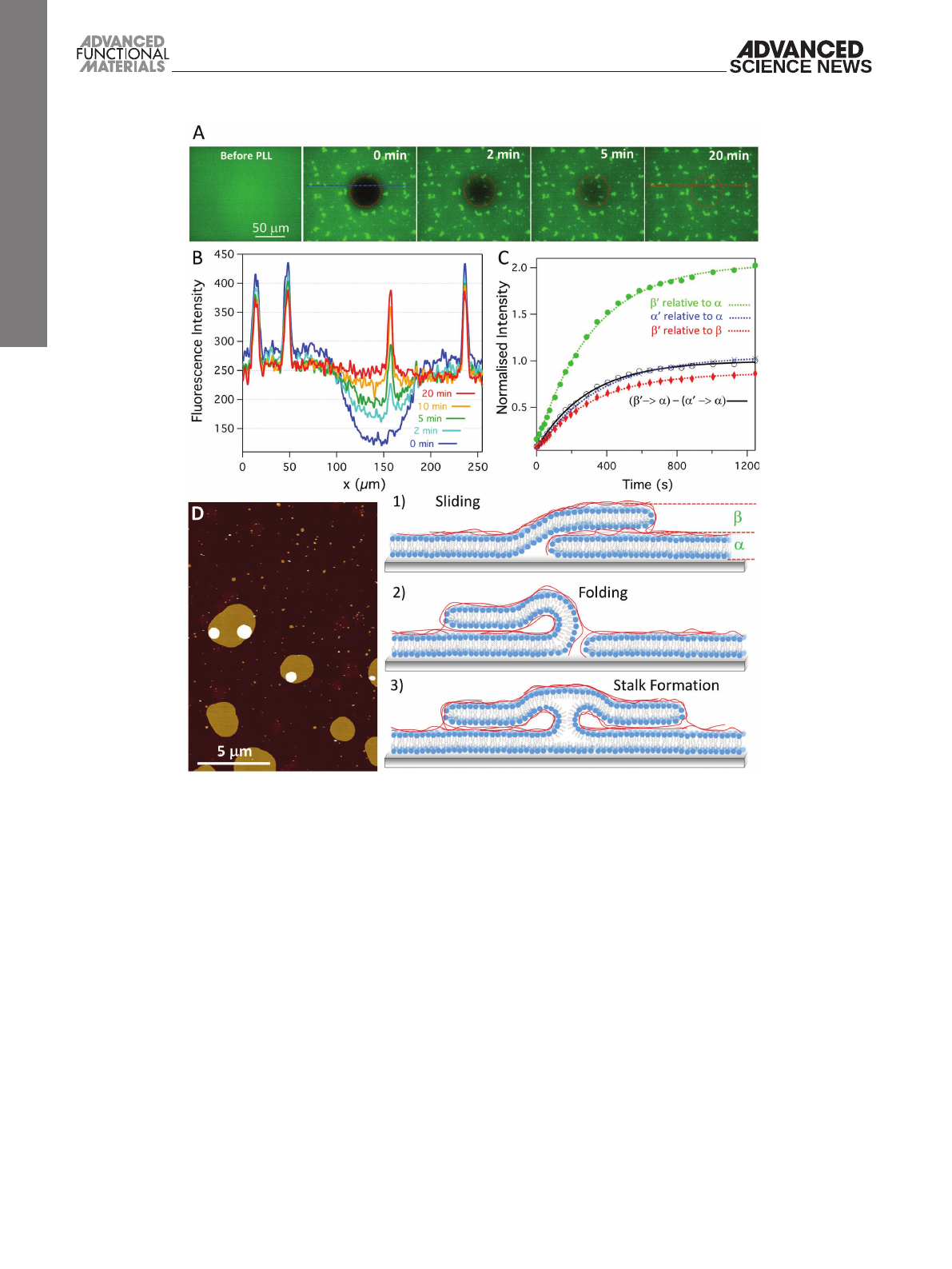
Figure 2A displays fluorescence images of a single solid
supported bilayer membrane (doped with a fluorescent lipid
analog) on which a layer of PLL was adsorbed. Before PLL, a
homogenous membrane is present as evidenced by a uniform
fluorescence intensity across the field of view. After addition
of PLL, discrete regions with twofold fluorescent intensity are
evident (Figure 2A,B). These patches are also seen by AFM
(Figure 2D), and the height and fluorescence intensity are con-
sistent with the formation of two lipid bilayer membranes on
top of each other. These patches appear before adding any addi-
tional vesicles. Furthermore, before addition of PLL, no vesicles
were detected on top of the base lipid bilayer (Figure 2A). This
suggests that the lipid required to form the double membrane
patches is extracted from the base bilayer. To test if the second
“top” membrane patches are continuous with the first mem-
brane beneath, we performed fluorescence recovery after photo-
bleaching (FRAP) experiments (Figure 2A). Importantly, after
photobleaching both the base lipid membrane and the double
membrane patches show full fluorescent recovery with nearly
identical rates (Figure 2B,C). Analysis of the intensities of
the bleached double patches relative to the nonbleached base
bilayer (Figure 2C) shows recovery to 197% after 20 min. This
indicates that fluorescently labeled lipids rapidly diffuse into
both membranes in the double membrane patches. Recovery
to the double membrane patches can be analyzed in isolation
by subtracting fluorescence intensity from the base membrane.
Performing this subtraction (Figure 2C, black line) results in
a fluorescent recovery that is almost indistinguishable from
that of the base bilayer (Figure 2C, blue line). The lateral dif-
fusion constants of 0.7 ± 0.2 µm2 s−1 and 0.6 ± 0.1 µm2 s−1 for
the double patches and base bilayer, respectively, indicate that
diffusion into double membrane patches is indistinguishable
from the lateral diffusion throughout the base bilayer. Such
www.afm-journal.dewww.advancedsciencenews.com
Adv. Funct. Mater. 2017, 27, 1606265
Figure 1. Schematic representation of the method used to create lipid–protein multilayers on gold electrodes. (1) Negatively charged vesicles are added
to the gold electrode that has a 6-mercaptohexanol and mercapto-(ethylene-oxy)3-cholesteryl (at an ≈3:2 ratio) self-assembled monolayer aiding the
formation of the first planar lipid bilayer. (2) Poly-l-lysine is bound to the surface of the lipid bilayer creating a net positive surface charge. (3) More
negatively charged vesicles are added, which rupture on the poly-l-lysine-coated membrane to form a double membrane. (4) Adsorption of poly-l-lysine
to the double membrane. (5) Steps (3) and (4) are repeated n times to create n additional stacked bilayers.
full paper
© 2017 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
wileyonlinelibrary.com
1606265 (3 of 9)
similar rates of diffusion imply that the two membranes in
the double membrane patches are interconnected via a lipid
phase. Similar patches of membrane stacks that are connected
to an underlying bilayer have previously been observed after the
addition of lipopolysaccharide[14] and, very recently, long chain
polyamines.[15]
As illustrated by the cartoons in Figure 2, there are a number
of possible configurations that allow rapid perpendicular diffu-
sion between the base and secondary membranes. Sliding or
folding structures may be induced by PLL in regions where
the base bilayer had a defect. Alternatively, rapid lipid transfer
between vesicles or from vesicles to a supported lipid bilayer
has previously been reported when the lipids adopt a so-called
stalk structure.[16,17] Typically, these stalks, which may be stabi-
lized by PLL, require two proximal leaflets to form a local neck-
like connection, resulting in a rapid lipid exchange between the
opposing membranes through lipid diffusion in the connected
leaflets.[18]
The formation of double membrane patches was monitored
in situ by AFM, which showed, upon addition of PLL, the
immediate formation (within tens of seconds; see Figure S1
in the Supporting Information) of double membrane patches
with clearly distinct sizes (Figure 2D): those with large diam-
eters of 500–5000 nm (3% by number) and those with diame-
ters averaging 90 ± 40 nm (the remaining 97%). The size of the
smaller patches remained stable over time, but larger patches
www.afm-journal.de www.advancedsciencenews.com
Adv. Funct. Mater. 2017, 27, 1606265
Figure 2. A) Series of fluorescence microscopy images before and after addition of PLL. After addition of PLL, fluorescence recovery is shown after
photo bleaching a central circular spot of a POPC/POPG 1:1 bilayer (0.5% TxRed DHPE). Note that the patches with double fluorescent intensity are
formed only after incubation with PLL. B) Intensity profiles across dashed lines in (A) at different time points. C) Plots of normalized fluorescence inten-
sity change where
α
and
β
denote intensity of single and double-bilayer patches, respectively, and prime (
α
′ and
β
″) the intensity within the bleached
area. The intensities of the bleached areas are normalized against either the base bilayer (
α
) or the double membrane patch (
β
), as indicated. The
black line represents the difference between “
β
′relative to
β
” and “
α
′relative to
α
”, as further explained in the text. D) AFM image of POPC/POPG
(1:1) bilayer after incubation with PLL (z-scale = 16.0 nm). The cartoon illustrations show the possible configuration that can allow diffusion between
multiple membranes: (1) sliding, (2) folding, or (3) stalk structures. Experiments were performed in 5 × 10−3 m MOPS, pH 7.0, 22 °C.

full paper
© 2017 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim wileyonlinelibrary.com (4 of 9) 1606265
were observed to grow across the bilayer asymmetrically (i.e.,
in one direction only; see Figure S1 in the Supporting Informa-
tion). Although the resolution of the AFM is not sufficient to
confirm this, we speculate that the larger patches are created by
sliding or folding of the membrane, consistent with the growth
in one direction only, and the smaller patches are due to the
stalk formation.
The further addition of negatively charged vesicles to
bilayers coated with PLL produces an almost complete second
bilayer. AFM indicates that the new second lipid bilayer mem-
brane seamlessly connects to the previous double membrane
patches (Figure S2, Supporting Information). It is thus pos-
sible that the structures shown schematically in Figure 2 are
maintained when multilayers are formed. Further addition of
PLL to this double membrane produces an even higher den-
sity of third bilayer patches, while closing up the second bilayer.
This process continues with each additional bilayer, potentially
providing diffusion routes between the whole multilayer.
2.2. Quinone Diffusion in Lipid Multilayers
Quinones are typical coenzymes in electron transport chains
and shuttle electrons between many membrane proteins that
are associated with energy conservation/transduction. The
FRAP experiments described above suggest that lipids rapidly
diffuse between lipid membranes in the multilayer membrane
patches. It is therefore likely that lipophilic quinones such as
ubiquinone-10 (UQ10) and menaquinone-7 (MQ7), which stay
in the hydrophobic region of the bilayer, should also transfer
between membranes. To test this hypothesis and to confirm
the presence of intermembrane connections, quinone-medi-
ated electron transfer across stacked lipid membranes was
investigated by cyclic voltammetry of lipid multilayers con-
taining MQ7 or UQ10. The multilayers were built up layer-
by-layer on mixed self-assembled monolayers (SAMs) on the
surface of gold electrodes. To adhere the first “base” lipid layer,
the membrane was tethered to the surface with previously
established methods.[19,20] In short, a mixed SAM was formed
on a gold electrode composed of mercapto-(ethylene-oxy)3-
cholesteryl (membrane tether, EO3C) and 6-mercaptohexanol
(spacer). The “base” lipid membrane was then formed via
self-assembly from lipid vesicles, where the cholesterol tether
induces rupture of the vesicle and allows the formation of a
continuous planar lipid membrane (the “base” lipid mem-
brane). Consecutive membrane layers were then formed by
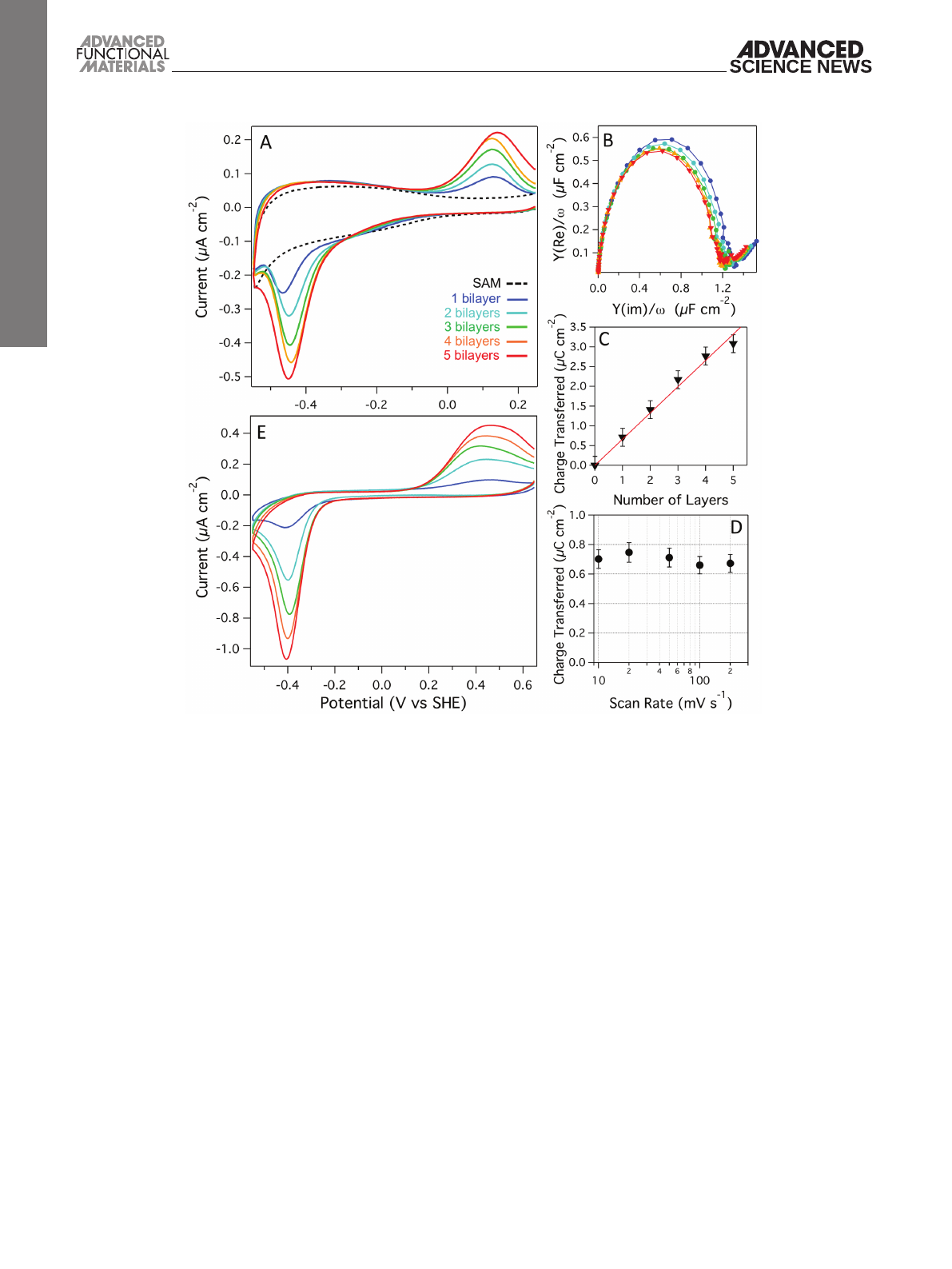
alternatively adsorbing PLL and vesicles (Figure 1). Figure 3A
shows a series of cyclic voltammograms (CVs) of the SAM
only, a 1:1 (w/w) POPC/POPG base membrane containing
1.5% (w/w) MQ7 and then each response after four cycles of
incubation with PLL followed by 3:1 POPC/POPG 1.5% (w/w)
MQ7 vesicles. These lipid compositions were chosen since we
have previously shown that this protocol creates additional
membranes with each PLL/vesicle incubation cycle on mica,
glass, and SiO2.[11] The CV of the first bilayer shows the MQ7
reduction (≈−0.45 V vs standard hydrogen electrode, SHE)
followed by oxidation (≈0.1 V vs SHE) peaks. The large peak
separation is partly caused by the thickness of the 6-mercap-
tohexanol layer in the SAM, reducing the electronic coupling
between electrode and MQ7. More importantly, however, it
is caused by the coupling of the electron transfer with pro-
tonation/deprotonation steps, which are slow due to the
lipid bilayer environment.[21] With each additional bilayer we
observe a linear increase in the current being transferred by
the MQ7 (i.e., increase in peak area), implying an increase in
the number of quinones that electrochemically interact with
the electrode.
The double-layer capacitance of the multilayer stacks
was estimated by electrochemical impedance spectroscopy
(Figure 3B). The formation of the first bilayer produced a
double-layer capacitance of ≈1.2 ± 0.1 µF cm−2 as estimated
from the diameter in the Cole–Cole plot. The capacitance is in
agreement with previous similar lipid membranes on the EO3C
system.[22] However, with each additional bilayer we observe
only small reductions in the double-layer capacitance, indi-
cating that the additional lipid bilayers have low resistance to
ions and thus likely contain defects.
CV analysis of the total charge transferred by MQ7
(Figure 3C) shows a linear increase for each additional lipid
membrane in the multilayer stack. An identically formed multi-
layer stack with UQ10 showed the same behavior (Figure S3,
Supporting Information). To determine if electron transfer was
limited by diffusion, CVs were performed on an eight-layered
system with scan rates varying between 10 and 200 mV s−1
(Figure 3D). The redox peak area shows no dependency on scan
rate, indicating that up to 200 mV s−1, UQ10 diffusion is not
limiting. There are no analytical solutions available to calcu-
late the diffusion in complex multilayered film systems such
as these, but it is clear that even in an eight-layered membrane
system, UQ10 in all layers is oxidized or reduced in less than a
second.
Although synthetic lipids offer a stable environment for
membrane proteins, activity of membrane proteins can be
dependent on the presence of particular annular lipids.[23] For
bacterial membrane enzymes, it can thus be beneficial to pro-
vide a more native lipid environment via the use of bacterial
lipid extracts. To study the layer-by-layer formation of multi-
layers composed of lipid extracts, we used E. coli “polar” lipid
extracts, which according to the supplier contain ≈23% phos-
phoglycerol (PG), 10% cardiolipin, and 67% phosphoethanola-
mine (w/w). The PG and cardiolipins give these membranes an
overall negative charge, as required for our LBL system with the
positively charged PLL. Figure 3E shows CVs at each stage after
incubating five cycles of E. coli polar extract vesicles containing
1% (w/w) UQ10. As with the POPC/POPG system, we observe
an increase in the quinone redox peaks with each addition of
vesicles/PLL. This suggests that, as long as the lipid mixture
contains sufficient negative charge, the LBL system can be
adapted to any lipid composition. It should be noted that E. coli
polar extract lipid also contains small amounts of ubiquinone-8,
native to E. coli.
From our results, the transfer of charge across multiple
membranes to the electrode via the reduction and oxidation
of the quinones is clear. What remains unclear is the mecha-
nism of how this process occurs. While AFM and fluores-
cence results suggest there are connections between adjacent
bilayers, allowing diffusion perpendicular to the plane of the
lipid membranes, there may be a number of other possible
www.afm-journal.dewww.advancedsciencenews.com
Adv. Funct. Mater. 2017, 27, 1606265
full paper
© 2017 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
wileyonlinelibrary.com
1606265 (5 of 9)
transfer routes such as quinone–quinone electron transfer,
where a quinone in one bilayer passes an electron to a quinone
in the adjacent bilayer, or quinone hopping across membranes.
To further test the mechanism of electron transport, a double
membrane system was prepared in which the quinones were
omitted in the base bilayer. In the latter system, the reduc-
tion and oxidation of the quinones is still observed (Figure S4,
Supporting Information) confirming that the quinones are
able to diffuse between the membranes layers. Spontaneous
lipid transfer between membranes is very slow (timescale of
hours).[24,25] To test if the same applies to UQ10 transfer, a
single-membrane layer was formed without UQ10 and incu-
bated with a liposome solution containing UQ10. As expected,
no UQ10 was observed to transfer from the liposomes in solu-
tion to the membrane on the surface (Figure S5, Supporting
Information). Finally, to exclude direct quinol to quinone elec-
tron transfer across membranes, single bilayers containing
UQ10 were formed and incubated with liposomes also con-
taining UQ10. The addition of liposomes did not alter the
UQ10 signals, confirming that electrons are not passed on
from the membrane on the surface to liposomes in solution
(Figure S5, Supporting Information). Taken together, these
results strongly suggest that the primary mechanism is diffu-
sion of quinols through the lipid interconnections between the
membrane stacks.
For a given molecule to transfer between two adjacent
membranes, it needs to diffuse a certain distance to the
nearest membrane connection. As determined from AFM
images of double membrane patches on the base bilayer, the
average nearest neighbor distance between patches is 470 nm
(SD ± 250 nm). Assuming a quinone or lipid molecule would
need to travel on average half this distance to reach an inter-
membrane connection, we can roughly estimate the time
taken (t) to diffuse this distance (x) using the 2D diffusion
equation: t = x2/4D, where D is the diffusion constant. Given
the lateral diffusion constant of 2 µm2 s−1 for UQ10[26] it would
take ≈0.01 s to diffuse to the next bilayer in the multilayer.
This timescale corresponds to the fact that it takes less than a
second to oxidize or reduce all UQ10 in an eight-layered mem-
brane system (see above).
www.afm-journal.de www.advancedsciencenews.com
Adv. Funct. Mater. 2017, 27, 1606265
Figure 3. A) Cyclic voltammograms, B) Cole–Cole plots, and C) total charge transferred during MQ7 reduction of POPC-POPG (1.5% MQ7)/PLL
multilayers. D) Charge transferred (reduction of UQ10) in CVs as a function of scan rate for a POPC-POPG (0.5% UQ10)/PLL multilayer with eight
layers. E) Cyclic voltammograms of E. coli polar lipid (1% UQ10)/PLL multilayers (color coding as in (A)). Experiments were performed in the absence
of O2 and at 10 mV s−1 scan rates, 20 × 10−3 m MOPS, 30 × 10−3 m Na2SO4, pH 7.4, 20 °C.
Loading more pages...