
Journal of Mathematical Biology (2021) 82:53
https://doi.org/10.1007/s00285-021-01596-0
Mathematical Biology
Separation of timescales for the seed bank diffusion and its
jump-diffusion limit
Jochen Blath1·Eugenio Buzzoni1·Adrián González Casanova2·
Maite Wilke Berenguer3
Received: 6 July 2018 / Revised: 1 October 2020 / Accepted: 27 October 2020 / Published online: 28 April 2021
© The Author(s) 2021
Abstract
We investigate scaling limits of the seed bank model when migration (to and from
the seed bank) is ‘slow’ compared to reproduction. This is motivated by models for
bacterial dormancy, where periods of dormancy can be orders of magnitude larger
than reproductive times. Speeding up time, we encounter a separation of timescales
phenomenon which leads to mathematically interesting observations, in particular
providing a prototypical example where the scaling limit of a continuous diffusion
will be a jump diffusion. For this situation, standard convergence results typically fail.
While such a situation could in principle be attacked by the sophisticated analytical
scheme of Kurtz (J Funct Anal 12:55–67, 1973), this will require significant technical
efforts. Instead, in our situation, we are able to identify and explicitly characterise a
well-definedlimitviadualityinasurprisinglynon-technicalway. Indeed, we show that
momentdualityisinasuitablesensestableunderpassagetothelimitandallowsadirect
and intuitive identification of the limiting semi-group while at the same time providing
a probabilistic interpretation of the model. We also obtain a general convergence
strategy for continuous-time Markov chains in a separation of timescales regime,
which is of independent interest.
Keywords Strong seed bank ·Two-island model ·Separation of timescales ·
Diffusion limits ·Jump-diffusion ·Duality
Mathematics Subject Classification Primary 60K35; Secondary 92D10
BMaite Wilke Berenguer
maite.wilkeberenguer@ruhr-uni-bochum.de
1Institut für Mathematik, Technische Universität Berlin, Berlin, Germany
2Instituto de Matemáticas, Universidad Nacional Autónoma de México, Mexico City, Mexico
3Fakultät für Mathematik, Ruhr-Universität Bochum, Bochum, Germany
123

53 Page 2 of 34 J. Blath et al.
1 Motivation and main results
In this extended introductory section, we first provide some background on the bio-
logical concept of dormancy and its relevance in particular in microbial communities.
This is followed by a short review of modelling approaches for dormancy in population
genetics, where we think that dormancy might be seen as an additional evolutionary
force, interacting with other forces such as genetic drift in complex ways. Since dor-
mancy periods vary over several orders of magnitude (depending on the underlying
species and environmental conditions), we aim for a systematic classification of rel-
evant timescales, leading to the distinction of three separate scaling regimes. While
the first two regimes have been modelled and analysed in population genetics before,
the last one, leading to a separation of timescales between genetic drift and dormancy
periods, is new, and completes the picture (at least on the level of ‘toy models’) of
modelling scenarios. Our results for this regime will be presented in this introduction
both for the forward-in time population model as well as for the dual genealogical
processes, leading to novel scaling limits, which are interesting also from a purely
mathematical perspective.
The proofs of these results can be found in Sects. 2and 3for the results going
backwards and forwards in time, respectively. We believe that our rather direct method
of proof to obtain and characterise these limits, making extensive use of duality for
Markov processes, can be applied in a variety of situations, so that in each section, we
first present the corresponding methodology in a general set-up and then discuss its
application to our concrete motivation.
Background on dormancy Dormancy is a complex trait that has developed indepen-
dently in many species across the tree of life and comes in many different guises.
Originally, theory for dormancy and the resulting seed banks has be developed in the
context of bet-hedging strategies for plants Cohen (1966). However, dormancy is also
a highly common trait in microbial communities, with important consequences for
their evolutionary, ecological and pathogenic properties.
Here, we define dormancy as the ability of (micro-) organisms to enter and leave
a state of vanishing metabolic activity. It has been observed for many habitats that at
any given time a large fraction of micro-organisms can be in such a dormant state. For
example, more than 80% of bacteria in soil are reported to be metabolically inactive,
forming large ‘seed banks’ comprised of dormant individuals, see Lennon and Jone
(2011). While dormancy seems to be an efficient and wide-spread strategy, e.g. to
withstand unfavourable environmental conditions, competitive pressure, or antibiotic
treatment, it is at the same time a costly trait whose maintenance involves energy and
a sophisticated ‘switching machinery’.
Dormancy also plays a role in various (human) diseases. So-called persister cells,
that may evade antibiotic treatment by remaining in a state of low activity, play a
major role in chronic infections, cf. Fisher et al. (2017), and individual cell dormancy
is linked to relapses in cancer, cf. Marx (2018), Endo and Inoue (2019).
In this paper, we will focus on microbial seed banks. Lennon and Jone (2011)
and Shoemaker and Lennon (2018) provide a broad overview of this rich and fas-
cinating field and serve as a motivation in the present paper. Given the relevance of
123

Separation of timescales for the seed bank diffusion… Page 3 of 34 53
biological systems exhibiting dormancy, investigating the mathematical implications
of dormancy in large populations seems to be a timely and interesting task.
Classification of the duration of dormancy: Known models and motivation for this
paper As indicated above, dormancy comes in many different forms, specific to the
involved species and environments. One variation lies in the duration of dormancy
periods: While in some microbial species dormancy periods last at most a few days,
others stay dormant for prolonged periods of time, and some, e.g. bacterial endospores,
have been reported to successfully resuscitate from dormancy after millions of years
(Shoemaker and Lennon 2018; Cano and Borucki 1995; Johnson et al. 2007; Morono
et al. 2020). The theoretical derivation and analysis of mathematical models may
help to identify, understand and classify the different effects of dormancy, on suitable
timescales, on the population dynamics and genealogical processes of the underlying
populations.
Hence, in this paper, we consider the consequences of dormancy and seed banks
in the framework of population genetics. More precisely, we are interested in the
interplay of dormancy and the classical evolutionary force of random genetic drift,in
particular with respect to its sensitivity to the duration of dormancy periods.
In a bi-allelic, haploid population that reproduces according to the Wright-Fisher
model, the frequency of a given allele converges to the Wright-Fisher diffusion,given
as the solution to
dZ(t)=Z(t)(1−Z(t))dB(t),
where (B(t))t≥0is a standard Brownian motion, if one measures time in the coalescent
timescale (alsoknown as the evolutionary timescale),i.e. on the order of the population
size as this tends to infinity. This diffusion is dual to the block-counting process of the
Kingman coalescent which in turn describes the genealogy of the population. These
objects serve as a reference for populations without dormancy and are widely studied
and applied in biology and mathematics alike. See e.g. Wakeley (2009) or Etheridge
(2011) for an overview. We will consider suitable extensions incorporating dormancy.
We propose to distinguish three regimes comparing the duration of dormancy peri-
ods to the coalescent timescale, i.e. the scale at which the random genetic drift acts.
1. Dormancy periods are small compared to the coalescent timescale.
In 2001, Kaj et al. (2001) introduced a model for dormancy in the following fashion:
insteadof alwayschoosing theancestorinthe preceding generations likein theWright-
Fisher model, individuals are allowed to choose an ancestor several generations in the
past. Their lineages thus ‘jump’ this number of generations and can be interpreted as
dormant during that time. If we denote by B≥1 the expected size of the ‘jump’, the
genealogy of the model converges on the coalescent timescale to a delayed Kingman
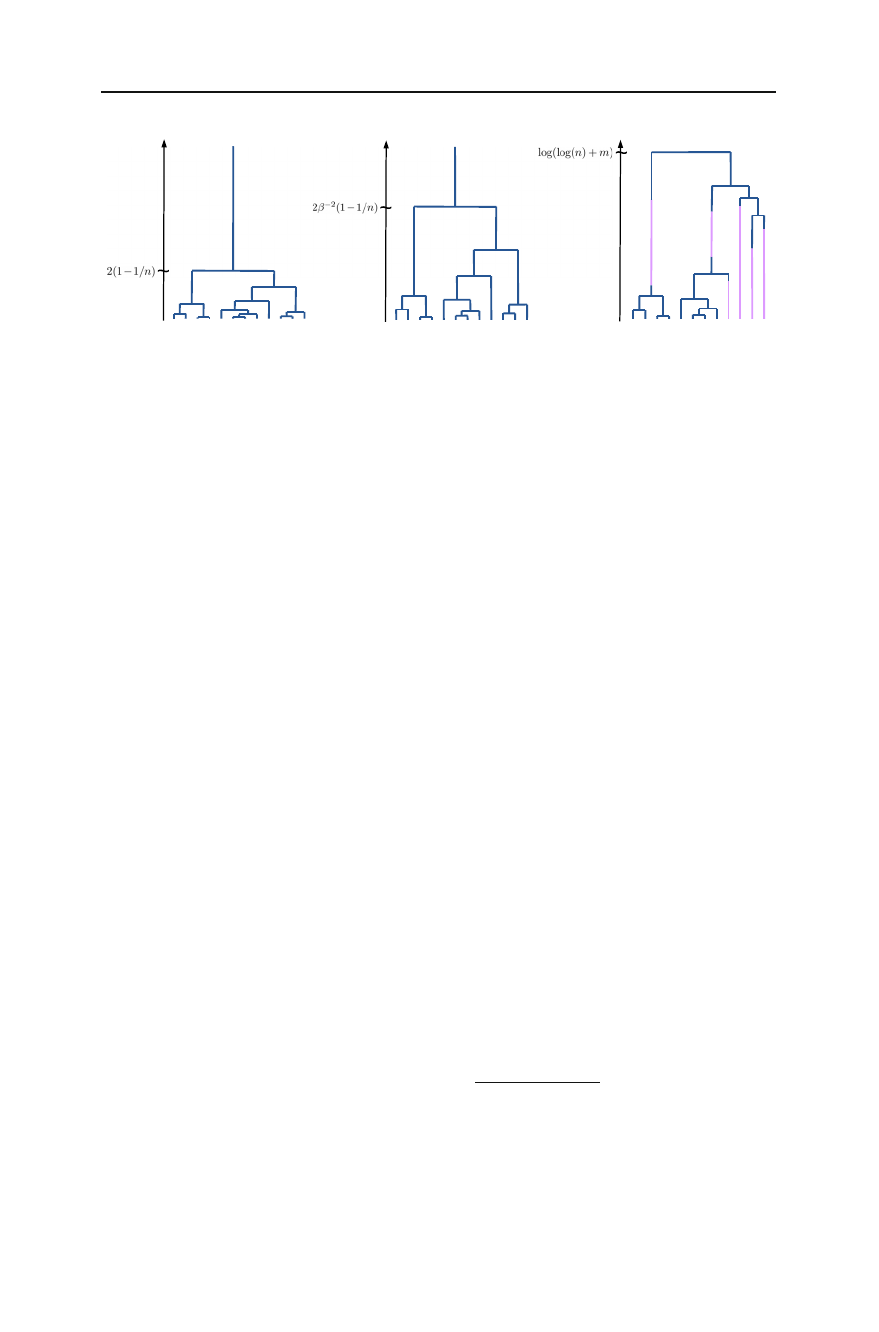
coalescent, depicted in Fig. 1b, where coalescences occur at rate β2, where β:= 1/B,
instead of at rate 1, cf. Kaj et al. (2001), Blath et al. (2013). This in turn is dual to the
delayed Wright-Fisher diffusion
d˜
Z(t)=β2˜
Z(t)(1−˜
Z(t))dB(t), (1)
123
53 Page 4 of 34 J. Blath et al.
(a) (b) (c)
Fig. 1 Typical realisations of athe Kingman coalescent, where lineages merge at rate 1 per pair, ba delayed
Kingman coalescent, where lineages merge at rate β2<1 per pair, and cthe seed bank coalescent, see Def.
1.2. In the seed bank coalescent there are two kinds of lines: blue lines are active lineages, while purple
lines are dormant lineages. The differences can be seen in the (asymptotic) expected time to the most recent
ancestor when started with a sample of n(active and mdormant) individuals given on the time-axis (colour
figure online)
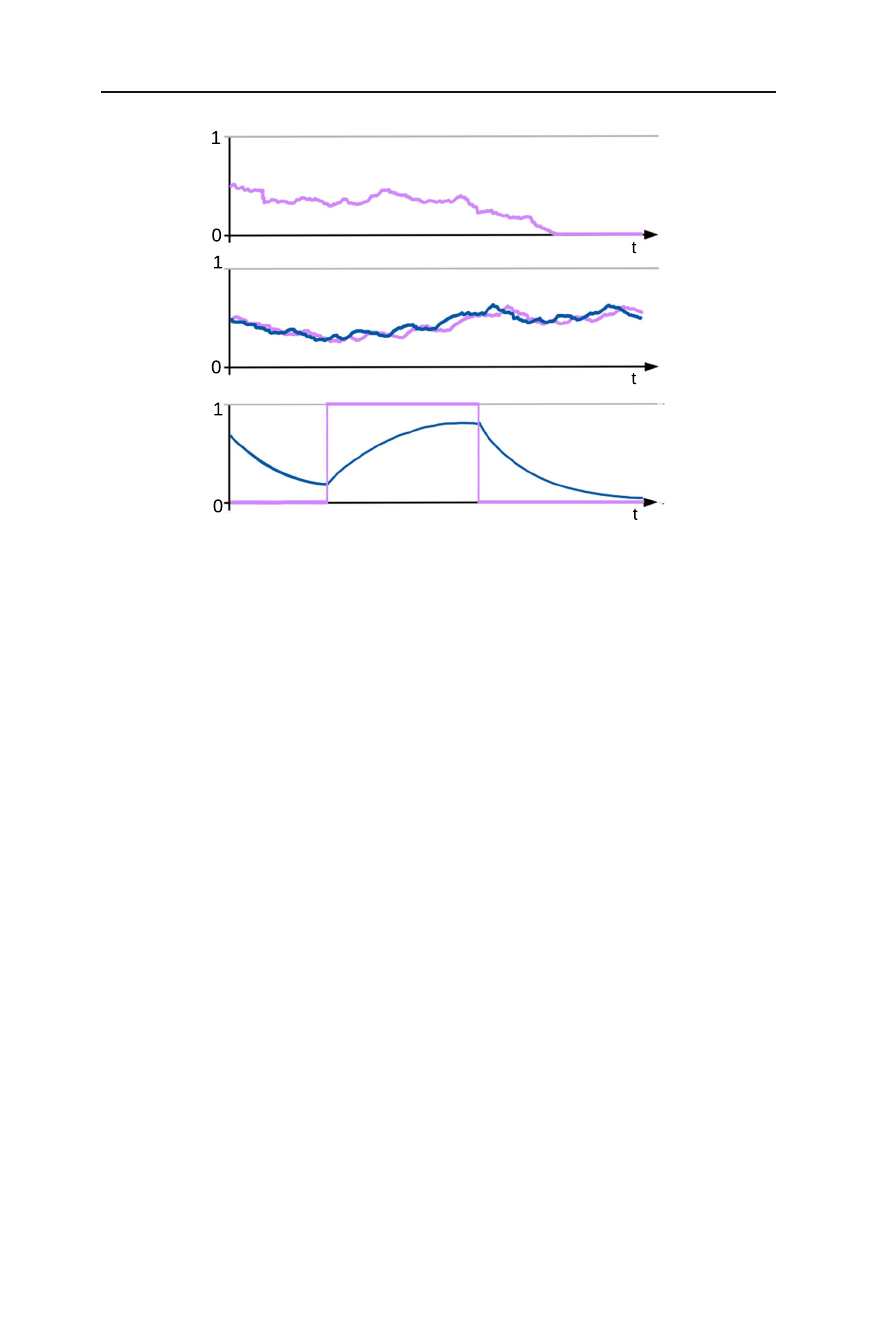
that again describes the frequency of a given allele in the population, cf. Fig. 2a. Note
that βdoes not depend on the population size, whence its qualitatively weak impact
on the coalescent timescale.
2. Dormancy periods on the order of the coalescent timescale
For microbial species, however, dormancy times can be much longer than just a
few ‘generations’, In this set-up, Lennon and Jone (2011) proposed a model based on
two reservoirs, the ‘active’ and the ‘dormant’ population, between which individuals
‘migrate/switch’ via initiation of and resuscitation from dormancy, at fixed rates. A
mathematical model for ‘spontaneous/stochastic’ switching (observed in nature under
stable environmental conditions, cf. Epstein 2009; Shoemaker and Lennon 2018), was
introduced and studied in Blath et al. (2016). This is reminiscent of the ‘two-island
model’ (Wright 1931; Moran 1959) with the notable difference of the absence of
reproduction on the second island.
If the size of the active and dormant population are proportional with the ratio
given by some K>0, the frequencies X(t)and Y(t)of a given allele in the active and
dormant population, respectively, when time is measured on the coalescent timescale,
are described by the seed bank diffusion, cf. Fig. 2b. This diffusion was first introduced
in Corollary 2.5 in Blath et al. (2016). The existence of a unique strong solution that
is Feller follows from Theorem 3.2 and Remark 3.2 in Shiga and Shimizu (1980), see
also Greven et al. (2020) for a more general seed bank diffusion.
Definition 1.1 (Seed bank diffusion)Let(B(t))t≥0be a standard Brownian motion and
c,Kfinite positive constants. The [0,1]2-valued continuous strong Markov process
(X(t), Y(t))t≥0given as the unique strong solution of the initial value problem
dX(t)=c(Y(t)−X(t))dt+√X(t)(1−X(t))dB(t),
dY(t)=Kc(X(t)−Y(t))dt,
(2)
with (X(0), Y(0)) =(x,y)∈[0,1]2, is called seed bank diffusion with parameters
c,K, starting at (x,y)∈[0,1]2.
123
Separation of timescales for the seed bank diffusion… Page 5 of 34 53
(a)
(b)
(c)
Fig. 2 Typical realisations of the trajectory of aa time-changed Wright-Fisher diffusion, where the time-
change is an effect of a weak seed bank, bthe seed bank diffusion, with the frequency of a given allele in
the active population displayed in blue and in the dormant population, in purple, cthe frequency process
(˜
X(t), ˜
Y(t)), using the same colour code (colour figure online)
The genealogy of such a population is given by the seed bank coalescent, introduced
in Definition 3.2 in Blath et al. (2016). Here, lineages can switch between an active and
a dormant state independently (hence ‘spontaneous’ switching) at a given rate c>0.
While the active lineages behave like the Kingman coalescent, dormant lineages are
prohibited from coalescing, as depicted in Fig. 1c.
That dormancy appears in such a prominent form in the coalescent and in the
diffusion and therefore is visible on the coalescent timescale is due to the underlying
scaling assumptions of the model. These imply that dormancy times are of the order of
the population size and therefore on the coalescent timescale. Here, many population
genetic quantities and statistics are affected in non-trivial ways, see Blath et al. (2015),
Blath et al. (2016) and Blath et al. (2020b) for a discussion of the scaling assumptions
and further extensions of the model. Since the seed bank here has a major qualitative
effect on both the diffusion and the coalescent, this is sometimes referred to as the
strong seed bank model.
As in the previous models, an important mathematical tool in our analysis will
be the formal duality relation between the seed bank diffusion (X(t), Y(t))t≥0and
the block-counting process of the seed bank coalescent (N(t), M(t))t≥0. Note that
the notion of a ‘block’ comes from the mathematical definition of a coalescent as a
partition-valued process. In thebiologicalcontext,the process could aswellbe denoted
the line-counting process, keeping track of the number of ancestral lines presents at
each time in the past.
Definition 1.2 (Block-counting process of the seed bank coalescent)LetE:= N0×
N0.Letc,K>0. We define (N(t), M(t))t≥0to be the continuous-time Markov chain
123
Loading more pages...