
ORIGINAL ARTICLE
Mono-specific forest plantations are valuable bat habitats:
implications for wind energy development
Sascha Buchholz
1,2
&Volker Kelm
3
&Simon J. Ghanem
3
Received: 6 March 2020 /Revised: 6 November 2020 /Accepted: 17 November 2020
#The Author(s) 2020
Abstract
Near-natural or semi-natural forests such as relatively undisturbed and old deciduous or mixed woodland are considered worth
protecting and ecologically valuable habitats for bat conservation. In contrast, mono-specific forest plantations are considered
ecologically less valuable; thus, decision-makers recommend these plantations as suitable locations for wind power stations and
therefore want to further expand wind turbines in these habitats. This is expected to have a strong negative impact on the
landscape because forests would be cleared for wind turbine pads and access roads and wind turbines rise above the trees with
adverse impacts for bats. Therefore, we argue that, in light of bat conservation, the suitability of forest plantations for wind energy
development is not, per se, warranted and that implications of wind power stations, even in mono-specific forest plantations,
should be assessed and evaluated. We conducted long-term bat activity monitoring and recorded bat echolocation calls above the
canopies of different forest sites (coniferous monoculture plantations and semi-natural mixed deciduous forests) in Germany and
compared different forest types in terms of species richness, total bat activity, activity of the three bat species groups and species
composition. Generalised linear models revealed that forest type and the amount of forest biotopes did not enhance bat activity.
Ordination showed that species composition was not affected by forest type, location and connectivity. Mono-specific forest
plantations can harbour a diverse bat fauna with high species activity and are, therefore, valuable bat habitats just as near-natural
or semi-natural woodlands are. Environmental impact assessment and mitigation measures are vital in all forest types before and
after planning for wind energy turbines. In particular, future planning and approval processes must consider the importance of
mono-specific forest plantations for bat species protection.
Keywords Chiroptera .Coniferous plantation .Environmental impact assessment .Renewable energy .Wind farm .Wind
power .Woodland
Introduction
Woodlands are vital for many bat species by providing habi-
tats, roosting sites and hunting grounds (e.g. Jantzen and
Fenton 2013; Kalda et al. 2015; Charbonnier et al. 2016a).
In general, the functional quality of woodlands is determined
by a number of landscape and habitat parameters such as
landscape connectivity and complexity (Boughey et al.
2011; Frey-Ehrenbold et al. 2013; Fuentes-Montemayor
et al. 2013). It has been widely shown that anthropogenic
impacts negatively influence the distribution of forest bat spe-
cies by affecting roosting and foraging habitats (frag-
mentation, Lesińskietal.2007; habitat loss, Crampton
and Barclay 1998; logging, Fukui et al. 2011). Furthermore,
negative changes in bat density and diversity are often
related to wind farms or wind turbines, respectively
(Kunz et al. 2007a,2007b;Hornetal.2008;Cryan
and Barclay 2009;Rydelletal.2010), which is of par-
ticular importance because many bat species are endangered at
global, national and regional levels (Mickleburgh et al. 2002;
Jones et al. 2009).
In Germany, renewable energies are forecasted to rise to
about 40–45% of the gross electricity consumption by 2025 to
*Sascha Buchholz
sascha.buchholz@tu-berlin.de
1
Department of Ecology, Technische Universität Berlin,
Rothenburgstr. 12, D-12165 Berlin, Germany
2
Berlin-Brandenburg Institute of Advanced Biodiversity Research
(BBIB), D-14195 Berlin, Germany
3
K&S Environmental Consult, Sanderstr. 28,
D-12047 Berlin, Germany
https://doi.org/10.1007/s10344-020-01440-8
/ Published online: 3 December 2020
European Journal of Wildlife Research (2021) 67: 1

fulfil climate protection goals and up to 80% by 2050, with
wind energy being the most important energy source
(Informationsportal Erneuerbare Energien 2020). Aside from
open landscapes, woodlands are considered potential loca-
tions for wind power stations. The German Federal Agency
of Nature Conservation (BFN 2011) estimates that about 14%
of existing German woodlands are suitable sites. In contrast to
near-natural or semi-natural woodlands, mono-specific forest
plantations are lower quality habitats for bats, and decision-
makers are recommending them for wind power stations (e.g.
MUGV Brandenburg 2011). Decision-makers argue that
mono-specific forest plantations—characterised by one tree
species (pine in NE-Germany) (Klimo et al. 2000)—are eco-
logically less valuable, as they emaciate the soil, are particu-
larly sensitive to windfall and pests and are less diverse in
plant and animal species (Klimo et al. 2000; MUGV
Brandenburg 2011). Therefore, wind energy development in
mono-specific forest plantations is seen as ecologically sound
and, thus, has become the focus of attention of wind power
operators (BFN 2011). The general consensus is there-
fore that the expansion of wind turbines in these plan-
tations should be promoted although landscape changes
are associated with this (for instance cleared forests for
wind turbine pads and access roads as, wind turbines
above the remaining forest patches).
In contrast, near-natural or semi-natural woodlands, such
as deciduous and mixed deciduous forests, are considered
worth protecting and ecologically valuable habitats, and sev-
eral studies prove their importance for bat conservation
(Kaňuch and Krištín 2005;Kaňuch and Celuch 2007; Zahn
et al. 2008; Kalda et al. 2015). This is especially true in terms
of wind energy development and its negative effects on bats
and bat habitats. It is known that higher collision rates may
occur due to higher bat activities above the canopy level
(Grunwald and Schäfer 2007; Brinkmann et al. 2011; Zahn
et al. 2014). Furthermore, indirect impacts, such as habitat
loss, can be expected through logging, as many bat species
strongly depend on tree holes as roosting sites (Meschede and
Heller 2000; Dietz et al. 2009).
To ensure environmentally sustainable energy production,
the implications of wind power stations in mono-specific for-
est plantations versus near-natural or semi-natural woodlands
should be assessed and evaluated. A number of studies
highlighted the adverse effects of wind power stations for bats
and bat habitats in woodlands (e.g. Brinkmann et al. 2011;
Hötker et al. 2006; Aschoff et al. 2006,Kunzetal.2007b),
but most studies focused on near-natural or semi-natural
woodlands while forest plantations have been nearly
neglected in Europe (but see Kirkpatrick et al. 2017a). This
is a drawback because the adverse effects of wind turbines
may even be stronger through the attraction effects of wind
turbines for bats (Cryan et al. 2014). We hypothesise that
mono-specific forest plantations can be valuable bat habitats
and that stand type is not necessarily a predictor of bat activity
as long as appropriate features such as tree holes, food avail-
ability and beneficial habitat structure or connectivity, respec-
tively, are provided at the landscape and habitat level
(Charbonnier et al. 2016b).
Methods
Field methods
We recorded bat echolocation calls with the batcorder system
(Batcorder—ecoObs, Nuremberg, Germany) above the cano-
pies of 24 different forest sites in Germany (Appendices Fig. 3
and Table 3). All sites were in proposed wind farms in a
landscape dominated by different forest types. Bat activity
can be highly variable with the daily variation of activity pat-
terns being as high as the variation between different sites
(Mathews et al. 2016, pers. observation). There are several
guidelines recommending minimum buffer zones of 200 m
to habitats of importance to bats (Rodrigues et al. 2015;
Kelm et al. 2014). Consequently, we defined a 200-m radius
for each location around the recording device to assess the
environmental variables using QGIS 3.4 (Quantum GIS
Development Team 2018 - © GeoBasis-DE/LGB, dl-de/by-
2-0). In this radius, we calculated the amount of the forest
biotopes (in percent) within each site and evaluated the avail-
able forest type and assigned it as a coniferous plantation
(consisted of a single pine species (Pinus sylvestris)) or mixed
deciduous forest (if at least 50% of the forest cover belonged
to this category) (Table 1). All forest patches showed a similar
age structure for around 50 years (± 20 years). To assess the
connectivity, we measured all linear landscape elements, e.g.
forest edges, hedges, roads or streams. We measured the dis-
tance (in m) from the recording device to the nearest forest
edge that marked a change in habitat type (roads or streams
were not considered) or water body. We defined water bodies
as those with standing water and a minimum size of 8 × 8 m.
At each site, we recorded calls for 57–183 entire nights
(sunset to sunrise) between April and November from 2011
to 2018. To ensure comparability, all surveys were conducted
on mild nights (> 10 °C ambient temperature), without rain
and low wind speeds (visual assessment with ≤small branches
moving, which according to the Beaufort scale is a maximum
speed of 3.4–5.4 m/s). We detected the bat passes above the
canopy with a batcorder fixed in a weatherproof box that was
attached on a pole protruding the treetops about 1.5 m. The
data transmission was facilitated via a GSM module, and the
power supply was provided by a solar panel. All batcorders
recorded the full frequency spectrum between 16 and
150 kHz in real time with a sampling rate of 500 kHz. We
used a threshold of 27 dB and a pre-trigger of 400 ms
for the recordings.
1 Page 2 of 12 Eur J Wildl Res (2021) 67: 1

We analysed the ultrasound recordings with the batIdent
and bcAnalyze software (ecoObs, Nuremberg, Germany) to
organise and identify species-specific echolocation calls. This
software automatically measures the primary sound parame-
ters of the recorded echolocation calls and calculates a proba-
bility value that a call is from a specific bat species. The
software identifies bat echolocation calls from simultaneous
recordings of up to three individuals per sequence (Runkel
and Marckmann 2009). After the automatic analysis, we man-
ually validated all echolocation calls for misidentifications
and, when possible, identified additional calls following the
criteria of species identification described in Hammer and
Zahn (2009)andSkiba(2003). Where the recording quality
allowed, we identified the following bats to species level:
Barbastella barbastellus,Eptesicus serotinus,Myotis
daubentonii,Myotis myotis,Myotis nattereri,Nyctalus
leisleri,Nyctalus noctula,Pipistrellus nathusii,Pipistrellus
pipistrellus,Pipistrellus pygmaeus. As some bat species are
acoustically nearly indistinguishable, we combined the re-
cordings of Myotis brandtii and Myotis mystacinus to the spe-
cies group Mbra and the species Plecotus auritus and Plecotus
austriacus to the species group Plecotus. Furthermore, we
included all Myotis-like, Nyctalus-like, Pipistrellus-like and
Plecotus-like calls in the categories Myotis sp., Nyctaloid,
Pipistrelloid and Plecotus sp. For testing environmental ef-
fects on bat species composition and activity, we first classi-
fied all bat species following their collision risk at wind tur-
bines as well as their foraging and echolocation behaviour to
one of the three groups, Nyctaloid group (high collision risk—
open space forager), Pipistrelloid group (medium collision
risk—edge space forager) and Myotis/Plecotus group (low
collision risk—narrow space forager). For detailed informa-
tion about classifying bats in groups and guilds, see Denzinger
and Schnitzler (2013). To describe bat activity, we counted the
number of bat passes for each of the different species or
groups. A ‘bat pass’was defined as a sequence of at
least two echolocation pulses of a passing bat (Thomas
and West 1989). Therefore, we assume that the number
of bat passes is highly correlated with bat activity. In
addition to the already high reliability and precision of
the batcorder system, we professionally calibrated all
used microphones prior to their field use to warrant the same
detectability under similar conditions. Consequently, all zero
observations were regarded statistically as true absences of
bats in a certain range.
Data analysis
The data analysis was done using the free software package
R-3.4.4 (R Core Team 2019). To test, if environmental vari-
ables(explanatoryvariables:amountofforestpatches,con-
nectivity, distance to forest edge, distance to next water body,
forest type) had a significant effect (alpha = P< 0.05) on total
bat activity, the species group activity and species numbers as
response variables, we performed generalised linear models
(GLM). Furthermore, we used the number of recording days
as an offset variable to correct for different recording intensi-
ties. To avoid multicollinearity, we only included environ-
mental parameters with correlations of |r| < 0.7. To compen-
sate for the detected overdispersion, the standard errors were
corrected using a quasi-Poisson GLM (Crawley 2007;Zuur
et al. 2009). The most appropriate models were stepwise
(backward) determined by analysis of deviance (Zuur et al.
2009). The residual deviance was used as a goodness-of-fit
measure by calculating the pseudo-R
2
(Dobson 2002). Direct
comparisons of categorical variable ‘forest type’were con-
ducted with post hoc Tukey’s pairwise comparisons using R
package LSMEANS.
In order to detect groupings of sample sites and the impact
of environmental variables on species composition, bat fre-
quencies were subjected to non-metric multidimensional scal-
ing (NMDS) using VEGAN and MASS packages in R. For
ordination, the frequencies of each species were square root
transformed. NMDS is an iterative ordination method that
places samples in a k-dimensional space using the ranked
distances between them (McCune and Grace 2002). The ordi-
nation was based on the Bray-Curtis dissimilarity matrix. In
search of a stable solution, a maximum of 100 random starts
was used. We applied Kruskal’s stress formula multiplied by
100, as given in McCune and Grace (2002). After two trials,
two convergent solutions were found for a three-dimensional
model. Afterwards, environmental data was fitted onto the
ordination. The significance of the explanatory power of each
variable was evaluated with a Monte-Carlo randomisation test
(10,000 permutations) (Manly 1997) using R function
‘envfit’.
Table 1 Description of environmental variables
Variable name Scale Explanation
Amount of forest patches Ratio % in a 200-m radius around the study plot
Connectivity Metric Linear landscape elements in a 200-m radius, e.g. forest edges, hedges, streams, roads
Distance to the forest edge Metric Is the distance in metres from the study plot to the nearest forest edge
Distance to the next water body Metric Is the distance in metres from the study plot to the nearest water body (minimum pond size)
Forest type Categorical Coniferous plantation, mixed deciduous
Page 3 of 12 1Eur J Wildl Res (2021) 67: 1

Results
In all, we detected 10 bat species (Appendix Table 3). Most
frequent were Nyctalus noctula,Pipistrellus pipistrellus,
Pipistrellus pygmaeus and Pipistrellus nathusii that occurred
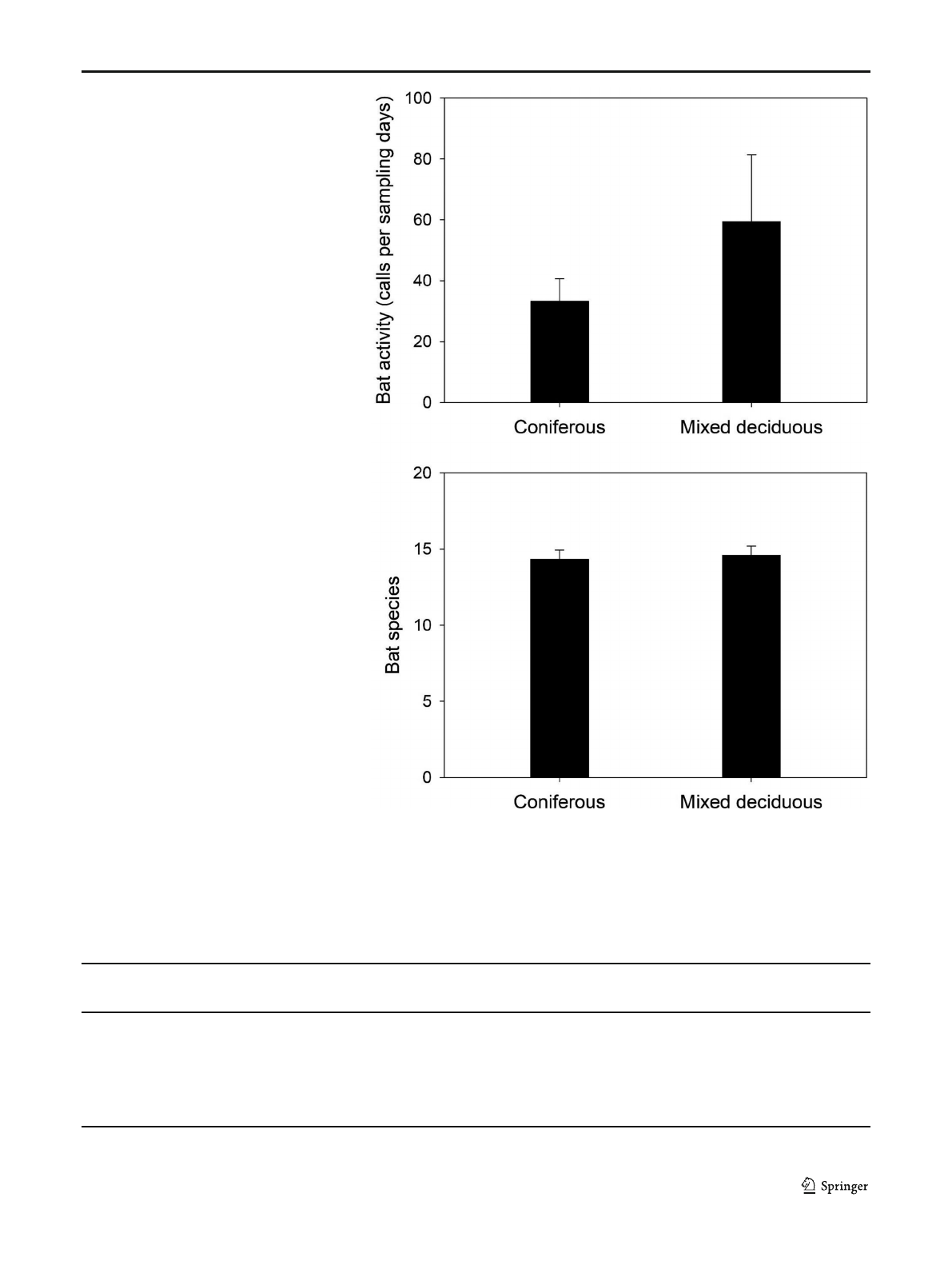
at all study sites. Generalised linear models revealed that total
bat activity (total bat passes) (Chi
2
= 1.56, P= 0.20, GLM)
and number of bat species (Chi
2
=0.65, P= 0.42, GLM) in
coniferous plantations and mixed deciduous forests did not
differ significantly (Fig. 1). Moreover, forest type did not sig-
nificantly affect the activity level of the three species groups
(Table 2), Myotis/Plecotus group (Chi
2
=0.48, P=0.49,
GLM), Nyctaloid group (Chi
2
= 0.83, P= 0.36, GLM) and
Pipistrelloid group (Chi
2
=2.82,P= 0.09, GLM). Bat activity
level in general (Chi
2
=3.87,P= 0.05, GLM) and the activity
level of the Nyctaloid (Chi
2
=3.72, P=0.05, GLM) and
Pipistrelloid groups (Chi
2
=3.07, P= 0.08, GLM) increased
with closer distance to the next water body as a statistical
trend. Also, as a statistical trend, the amount of forest
patches negatively affected total bat activity (Chi
2
=
3.40, P=0.07, GLM) and Nyctaloid group activity
(Chi
2
=3.34, P= 0.07, GLM) while Myotis/Plecotus spe-
cies responded positively (Chi
2
=3.08, P=0.08, GLM).
All other environmental variables had no impact on the
activity levels of the groups or on the number of en-
countered species.
The NMDS based on the bat species activity level revealed
no significant effects of the environmental variables (Fig. 2).
None of the variables (connectivity, distance to forest edge,
distance to next water body, forest or forest type) af-
fected species groups (amount of forest patches: F=0.1,
R
2
=0.01, P= 0.96; connectivity: F=0.4, R
2
=0.02, P=
0.73; distance to forest edge: F=0.4, R
2
= 0.02, P=
0.75; distance to next water body: F=1.4, R
2
=0.06,
P= 0.25; forest type: F=2.1, R
2
=0.09, P=0.10; per-
mutational multivariate analysis of variance with 9999
permutations).
Discussion
Impact of forest type
It is well known that near-natural or semi-natural woodlands,
such as deciduous and mixed deciduous forests, are ecologi-
cally valuable bat habitats; thus, they are considered worth
protecting (e.g. Jantzen and Fenton 2013; Kalda et al. 2015;
Charbonnier et al. 2016a,2016b). However, we found no
significant differences in bat densities in mixed deciduous
forest and mono-specific forest plantations. Given that none
of the bat groups responded to forest type, we encourage a
change of thinking regarding the ecological value of mono-
specific forest plantations for bats. Even if forest plantations
are ecologically critical, they still provide a habitat with equal-
ly high bat activity as near-natural or semi-natural woodlands
(Pereira et al. 2016; da Silva et al. 2019). Moderate silvicul-
tural methods can also promote bat diversity (Kirkpatrick et al.
2017a), such as harvesting, which creates a patch mosaic with
different tree densities (Perry et al. 2007;Blakeyetal.2016;
Kirkpatrick et al. 2017b).
Kalda et al. (2015) found only a minor impact of forest type
on bat diversity. They concluded that natural and anthropo-
genic woodlands could be seen as a single land-cover type
regardless of the dominating tree species. In addition,
Glendell and Vaughan (2002) found forest plantations to be
suitable habitats for bats, as they showed that bats similarly
forage in semi-natural and plantation woodlands. Bats depend
on a variety of landscape and habitat properties, for example
forest edge structures for commuting and foraging (Hein et al.
2009;Kelmetal.2014;Morrisetal.2010; Jantzen and Fenton
2013), open patches (Loeb and O’Keefe 2006; Perry et al.
2007; Kirkpatrick et al. 2017b), diverse surrounding land-
scape compositions (Boughey et al. 2011; Frey-Ehrenbold
et al. 2013; Heim et al. 2018), water bodies (Morris et al.
2009; Heim et al. 2018) and roosting sites (Kühnert et al.
2016;Coronadoetal.2017; Dietz et al. 2018). However, all
these features potentially exist in all diverse, near-natural and
mono-specific forest plantations (Cruz et al. 2016;Kirkpatrick
et al. 2017a). According to Ruczyński et al. (2010) and Toth
et al. (2015), woodland bats may change their roost site
preferences and could, thus, occupy habitats previously
considered less suitable. Runkel (2009) observed that bats
generally use a wide range of forest habitats, but only a few
specialists succeed in niche partitioning in forests with a low
spacing between trees. Our results show that many open and
edge space species, such as Pipistrellus pipistrellus,
Pipistrellus nathusii,Eptesicus serotinus and Nyctalus
noctula, also had high activity in coniferous forestry zones
which adds to findings of Kirkpatrick et al. (2017b) for Sitka
spruce (Picea sitchensis) plantations.All these species are
negatively affected by wind energy development
(Brinkmann et al. 2011) and at risk from wind turbines
(Müller et al. 2013).
Differences in species composition naturally occur across
different regions; however, other drivers—such as landscape
composition, habitat structure and stand origin—are compa-
rable (Kalda et al. 2015). In our study, species composition
was not influenced by forest type, even though Pipistrellus
species usually show a preference for open patches in forests,
such as thinnings and windbreaks (Kirkpatrick et al. 2017b),
which are habitat conditions that rather occur in mixed decid-
uous forests. However, Pipistrellus pipistrellus is a generalist
species that can reach high densities in manifold landscapes
(Davidson-Watts et al. 2006). Moreover, Myotis daubentonii
can use open space landscapes (Kalda et al. 2015) but strongly
depends on ponds and lakes as primary foraging habitats
1 Page 4 of 12 Eur J Wildl Res (2021) 67: 1
(Glendell and Vaughan 2002; Dietz et al. 2009). In general,
Myotis species are known to select habitat structure over forest
type. Thus, bats of the Myotis genus are able to negotiate
cluttered woodland interiors (Glendell and Vaughan 2002).
Table 2 Regression results (quasi-Poisson GLM). Positive effects on dependent variables are expressed as ↑(for increasing) and negative ones are
expressed as ↓(for decreasing)
Response Amount of forest patches Connectivity Distance to the
forest edge
Distance to the
next water body
Forest type
Total bat passes all groups Chi
2
= 3.40, P=0.07(↓)Chi
2
= 0.40, P=0.53 Chi
2
=1.33,P=0.25 Chi
2
=3.87,P=0.05(↓)Chi
2
= 1.56, P=0.21
Species number Chi
2
= 1.53, P=0.22 Chi
2
= 0.26, P=0.61 Chi
2
=2.24,P=0.26 Chi
2
=1.26,P=0.26 Chi
2
= 0.65, P=0.42
Myotis/Plecotus group Chi
2
= 3.08, P=0.08(↑)Chi
2
= 0.20, P=0.65 Chi
2
=1.92,P=0.17 Chi
2
=1.32,P=0.25 Chi
2
= 0.48, P=0.49
Nyctaloid group Chi
2
= 3.34, P=0.07(↓)Chi
2
= 0.36, P=0.54 Chi
2
=1.29,P=0.26 Chi
2
=3.72,P=0.05(↓)Chi
2
= 0.83, P=0.36
Pipistrelloid group Chi
2
= 2.62, P=0.11 Chi
2
= 0.32, P=0.57 Chi
2
=1.05,P=0.31 Chi
2
=3.07,P=0.08(↓)Chi
2
= 2.82, P=0.09
Fig. 1 Bat activity and number of
bat species (mean and standard
error of the mean) in coniferous
plantations and mixed deciduous
forest did not differ significantly
between both forest types (bat
activity: Chi
2
=1.56,P=0.20,
GLM; number of bat species:
Chi
2
=0.65,P= 0.42, see
Table 2). For bat activity, addi-
tional post hoc Tukey’s pairwise
comparisons of categorical vari-
able ‘forest type’have been con-
ducted:
z.ratio = 1.23, P=0.22
Page 5 of 12 1Eur J Wildl Res (2021) 67: 1
Loading more pages...