
Suietal. Microb Cell Fact (2020) 19:198
https://doi.org/10.1186/s12934-020-01450-w
RESEARCH
Engineering cofactor metabolism
forimproved protein andglucoamylase
production inAspergillus niger
Yu‑fei Sui1,2, Tabea Schütze2 , Li‑ming Ouyang1, Hongzhong Lu3, Peng Liu1, Xianzun Xiao1, Jie Qi1,
Ying‑Ping Zhuang1* and Vera Meyer2*
Abstract
Background: Nicotinamide adenine dinucleotide phosphate (NADPH) is an important cofactor ensuring intracel‑
lular redox balance, anabolism and cell growth in all living systems. Our recent multi‑omics analyses of glucoamylase
(GlaA) biosynthesis in the filamentous fungal cell factory Aspergillus niger indicated that low availability of NADPH
might be a limiting factor for GlaA overproduction.
Results: We thus employed the Design‑Build‑Test‑Learn cycle for metabolic engineering to identify and prioritize
effective cofactor engineering strategies for GlaA overproduction. Based on available metabolomics and 13C meta‑
bolic flux analysis data, we individually overexpressed seven predicted genes encoding NADPH generation enzymes
under the control of the Tet‑on gene switch in two A. niger recipient strains, one carrying a single and one carrying
seven glaA gene copies, respectively, to test their individual effects on GlaA and total protein overproduction. Both
strains were selected to understand if a strong pull towards glaA biosynthesis (seven gene copies) mandates a higher
NADPH supply compared to the native condition (one gene copy). Detailed analysis of all 14 strains cultivated in
shake flask cultures uncovered that overexpression of the gsdA gene (glucose 6‑phosphate dehydrogenase), gndA
gene (6‑phosphogluconate dehydrogenase) and maeA gene (NADP‑dependent malic enzyme) supported GlaA
production on a subtle (10%) but significant level in the background strain carrying seven glaA gene copies. We thus
performed maltose‑limited chemostat cultures combining metabolome analysis for these three isolates to charac‑
terize metabolic‑level fluctuations caused by cofactor engineering. In these cultures, overexpression of either the
gndA or maeA gene increased the intracellular NADPH pool by 45% and 66%, and the yield of GlaA by 65% and 30%,
respectively. In contrast, overexpression of the gsdA gene had a negative effect on both total protein and glucoamyl‑
ase production.
Conclusions: This data suggests for the first time that increased NADPH availability can indeed underpin protein
and especially GlaA production in strains where a strong pull towards GlaA biosynthesis exists. This data also indicates
that the highest impact on GlaA production can be engineered on a genetic level by increasing the flux through
the pentose phosphate pathway (gndA gene) followed by engineering the flux through the reverse TCA cycle (maeA
gene). We thus propose that NADPH cofactor engineering is indeed a valid strategy for metabolic engineering of A.
© The Author(s) 2020. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and
the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material
in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material
is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the
permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creat iveco
mmons .org/licen ses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creat iveco mmons .org/publi cdoma in/
zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Open Access
Microbial Cell Factories
*Correspondence: [email protected].cn; vera.meyer@tu‑berlin.de
1 State Key Laboratory of Bioreactor Engineering, East China University
of Science and Technology, Shanghai 200237, People’s Republic of China
2 Chair of Applied and Molecular Microbiology, Institute of Biotechnology,
Technische Universität Berlin, Straße des 17. Juni 135, 10623 Berlin,
Germany
Full list of author information is available at the end of the article

Page 2 of 17
Suietal. Microb Cell Fact (2020) 19:198
Background
The filamentous fungus Aspergillus niger is one of the
main cell factories used nowadays in the industry for
homologous or heterologous protein production due
to its extraordinary ability for protein expression and
secretion [1–3]. The Design-Build-Test-Learn (DBTL)
cycle is an increasingly adopted systematic meta-
bolic engineering strategy to achieve the desired out-
come through reconstructing heterologous metabolic
pathways or rewiring native metabolic activities [4,
5]. Rational strain development of cell factories can
be improved by the iterative application of the DBTL
cycles, which not only contributes to the optimization of
biomanufacturing processes, it is also advantageous to
build a complete metabolic model of engineered cells to
deepen our understanding of cellular metabolism. Note-
worthy, the advance of genetic engineering has speeded
up the DBTL cycle of metabolic engineering [6]. For the
cell factory A. niger, several genetic approaches have
proven their potency to improve its enzyme producing
capability, including protein carrier approaches, tun-
able Tet-on driven gene expression, and morphology
engineering, to name but a few [1, 7–9]. However, the
impact of cofactor engineering, i.e., the rebalance of the
intracellular redox status, on protein production has not
been systematically studied in A. niger.
NADPH is a limiting factor for the biosynthesis of
amino acids that are the building blocks of proteins. For
instance, 3mol and 4mol of NADPH is required for pro-
ducing 1 mol of arginine and lysine, respectively [10].
Thus, adequate cytosolic NADPH supply is indispensa-
ble to maintain the intracellular redox balance and serves
as a driving force for efficient amino acid biosynthesis
[11]. NADPH also provides the main anabolic reducing
power for biomass growth, lipid formation, and also for
natural product biosynthesis [12]. Indeed, cofactor engi-
neering has been reported to improve productivities in
the bacterial cell factories Escherichia coli [13, 14], and
Corynebacterium glutamicum, as well as in the yeast cell
factory Yarrowia lipolytica [15]. Two common strategies
have mainly been employed to optimize the availabil-
ity of NADPH. One is to activate the enzyme activities
of NAD(H) kinases (EC 2.7.1.86, EC 2.7.1.23) which are
used to obtain NADPH or NADP + through phospho-
rylation of NADH and NAD + , respectively. The other is
to modulate the expression strength of typical NADPH
generating enzymes of the glycolytic pathway, the pen-
tose phosphate pathway or the citric acid cycle. These
include glucose-6-phosphate dehydrogenase (G6PDH),
6-phospho-gluconate dehydrogenase (6PGDH), NADP-
dependent isocitrate dehydrogenase (NADP-ICDH),
and NADP-dependent malic enzyme (NADP-ME) [16,
17]. Notably, heterologous protein expression in Pichia
pastoris and A. niger can be triggered through boosted
carbon flux to the pentose phosphate pathway (PPP), a
catabolic pathway also known to produce NADPH [18,
19]. This suggests that central carbon metabolism may
have evolved to ensure the production of cellular compo-
nents under the balance of energy production and con-
sumption [4]. In agreement, the metabolic flux through
the PPP increased by 15–26% compared to the parental
strains when GlaA was overproduced in A. niger [20] or
the enzyme amylase overproduced in A. oryzae [21].
In the past two decades, extensive studies have focused
on engineering a high flux through the PPP in E. coli,
C. glutamicum, A. nidulans, and A. niger [11, 22–25].
A block of the glycolytic pathway by down-regulating
the pgi gene encoding a phosphoglucose isomerase was
one successful strategy in C. glutamicum [11]. In order
to elevate the NADPH pool originating from the PPP
in A. niger, the gsdA gene (glucose 6-phosphate dehy-
drogenase), the gndA gene (6-phosphogluconate dehy-
drogenase) and the tktA gene (transketolase) were
individually overexpressed in A. niger. Strong overexpres-
sion of gndA led to a nine-fold increase in intracellular
NADPH concentration, while gsdA and tktA affected the
NADPH level only weakly [25]. However, any correlation
between the NADPH supply and enzyme overproduction
remained unclear.
Irrespective of the importance of the PPP for NADPH
regeneration, an efficient carbon economy is only guaran-
teed when the carbon flux enters the glycolytic pathway
(Embden-Meyerhoff-Parnass pathway, EMP) instead of the
PPP because the PPP releases one carbon as CO2 when oxi-
dizing 1mol of hexose. Takeno etal. [26] thus substituted
the endogenous NAD-dependent glyceraldehyde-3-phos-
phate dehydrogenase (GAPDH) in the EMP in C. glu-
tamicum with a heterologous NADP-dependent GAPDH,
leading to 2mol of NADPH generation instead of 2mol
NADH from 1mol of hexose. This genetic modification
niger to improve GlaA production, a strategy which is certainly also applicable to the rational design of other microbial
cell factories.
Keywords: Aspergillus niger, NADPH, Genetic engineering, CRISPR/Cas9, Tet‑on, Metabolic engineering, Chemostat,
Glucoamylase

Page 3 of 17
Suietal. Microb Cell Fact (2020) 19:198
provoked a substantial improvement in the yield of L-lysine
production by 70–120%. Similar strategies also have been
followed to overproduce ethanol in the yeast Saccharomy-
ces cerevisiae [27] or lycopene and ε-caprolactone in the
bacterium Clostridium acetobutylicum [28]. Likewise, cyto-
solic NADP-ME has been shown to positively affect lipid
accumulation in oleaginous fungi [29, 30].
As summarized above, a wealth of metabolomic and
fluxomic data in A. niger demonstrated that strains
adapted to protein overproducing conditions channel a
higher carbon flux through the PPP. However, cofactor
engineering has not been considered yet or performed
in A. niger to guide enzyme overproduction. We thus
mined our recently published genome-scale metabolic
network model (GSMM) developed for the A. niger pro-
tein producing reference strain CBS 513.88 [31]. This
iHL1210 model identified the involvement of NADPH
in 173 intracellular redox reactions in A. niger, includ-
ing 49 NADPH generating reactions [31]. Notably, the
GSMM did not predict any NADPH/NADP + shuttle in
the mitochondrial membrane, and we thus concluded
that any mitochondrial NADPH is unlikely to become
directly consumed by cytosolic amino acid biosynthe-
sis. Overall, the GSMM predicted that seven potential
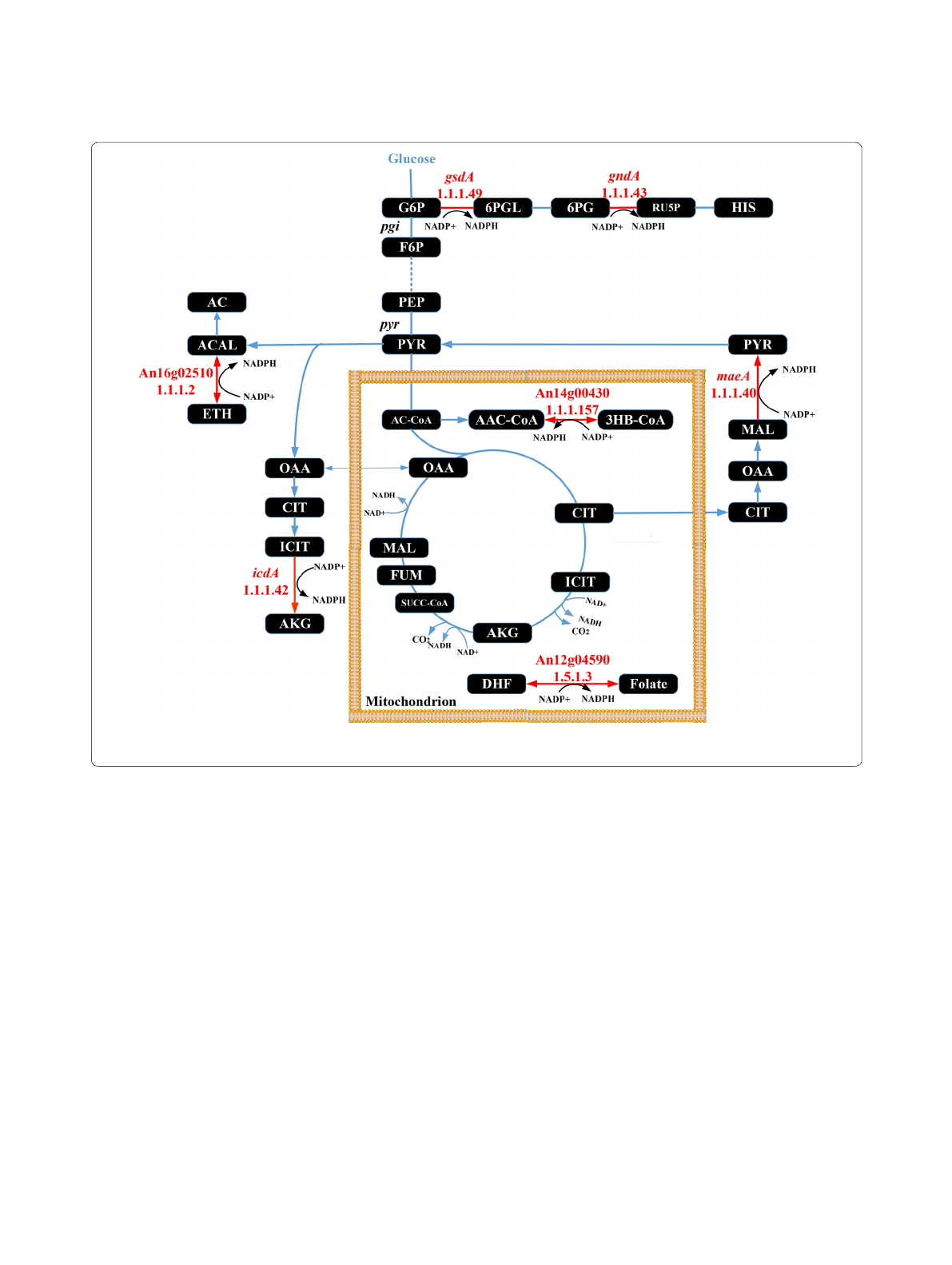
NADPH generating enzymes are of importance for GlaA
production in A. niger (Table1, Fig.1): two enzymes of
the cytosolic PPP (glucose-6-phosphate dehydrogenase,
G6PDH; 6-phosphogluconate dehydrogenase 6PGDH),
two cytosolic NADP-dependent enzymes (NADP-ICDH
and NADP-ME) and three uncharacterized open read-
ing frames (An12g04590, An14g00430, An16g02510).
An12g04590, An14g00430 show high homology to
NADP + oxidoreductases, and An16g02510 displays
homology to alcohol dehydrogenases. In order to evalu-
ate whether the model prediction is strain-dependent,
we individually overexpressed all seven candidate genes
in two A. niger host strains. Strain AB4.1 produces native
levels of GlaA as it carries one glaA gene copy, and strain
B36 is a derivative thereof, carrying seven glaA gene
copies and is thus a high-yield GlaA producing strain
[32]. All 14 strains were first investigated in shake flask-
level cultivations. Based on the data gained, three engi-
neered strains were selected for chemostat cultivations
to decipher the association among genetic perturbation,
NADPH availability, and GlaA production in A. niger.
Results
Strain generation using CRISPR/Cas9 technology
andthesynthetic Tet‑on gene switch
In order to compare the effect of the seven selected genes
on GlaA production in an A. niger strain carrying one
glaA (AB4.1) or seven glaA (B36) gene copies, we first
had to ensure that the introduced genetic modifications
would allow us to directly compare the observed phe-
notypes. This required that the introduced genes would
be under the same genetic control and furthermore
introduced at the same genomic locus in both recipient
strains. We thus decided to integrate an additional copy
of all candidate genes under the control of the strong
and tunable Tet-on gene switch into the pyrG locus of
A. niger. This gene switch is inducible by the addition of
doxycycline (DOX) to the culture medium, is tight in the
absence of DOX and metabolism-independent in A. niger
[33]. It has furthermore been shown to strongly induce
gene expression up to levels above the glucoamylase
gene, which is one of the highest expressed genes in A.
niger [33–35].
Strain AB4.1 is a uridine-auxotroph due to a defective
pyrG gene, a locus that is perfectly suited for gene tar-
geting and screening purposes. The introduction of an
Table 1 GSMM-predicted NADPH producing reactions inA. niger
[m] reactions in the mitochondrion; Rn, reaction. All reactions listed here were predicted in Lu etal. [31]
Rn
name Rn description Formula Gene
R25 Glucose 6‑phosphate‑dehydrogenase (gsdA) G6P + NADP = > D6PGL + NADPH + H An02g12140
R27 Phosphogluconate dehydrogenase (gndA) 6PGC + NADP = > Ru5P + CO2 + NADP An11g02040
R36 Isocitrate dehydrogenase (icdA) (NADP +) ICIT[m] + NADP[m] = > AKG[m] + CO2[m] + NADPH[m] An02g12430
R38 Isocitrate dehydrogenase (NADP +) ICIT + NADP = > AKG + CO2 + NADPH An02g12430
R55 Malic enzyme
(NADP‑specific) (maeA)MAL + NADP = > PYR + CO2 + NADPH An05g00930
R57 Malic enzyme
(NADP‑specific) MAL[m] + NADP [m] = > PYR
[m] + CO2[m] + NADPH [m] An05g00930
R110 (S)‑3‑Hydroxybutanoyl‑CoA: NADP + oxidoreductase 3HBCoA[m] + NADP [m] < = > AACCoA[m] + NADPH [m] +H[m] An14g00430
R125 dihydrofolate:NADP + oxidoreductase NADP [m] + DHF[m] < = > NADPH [m] + FOLATE[m] An12g04590
R190 Alcohol dehydrogenase ETH + NADP < = > ACAL + NADPH + H An16g02510
Page 4 of 17
Suietal. Microb Cell Fact (2020) 19:198
intact pyrG copy at this locus occurs efficiently and con-
fers uridine prototrophy in A. niger [36]. However, strain
B36 does carry an intact pyrG gene [37]. We thus first
mutated the pyrG locus in this strain in order to apply the
same gene targeting strategy for all seven genes in both
recipient strains. We edited the pyrG gene in B36 by fol-
lowing a CRISPR/Cas9 strategy that employed ribonu-
cleoprotein particles. This approach was first published
for the penicillin producer Penicillium chrysogenum [38]
and has later been successfully established in other fun-
gal cell factories [39]. As explained in detail in Additional
file1: Fig. S1, this approach enabled us to obtain a deriva-
tive of B36, strain YS20.2, which carries a 195bp deletion
within the pyrG ORF and is therefore unable to grow on
medium lacking uridine or uracil. Both recipient strains,
AB4.1 and YS20.2, were eventually used to integrate Tet-
on driven candidate genes at the pyrG locus (for details
see Materials and Methods and Additional file 1: Fig.
S2). Respective genetic modifications were proven by
PCR and Southern blot analyses (Additional file1: Figs.
S3, S5, S6, S7). All 14 strains obtained are summarized in
Table3. We finally also decided to delete the native ORFs
of An14g00430 and An16g02510 in their respective Tet-
on driven overexpression strains in order to analyze their
deletion phenotypes.
The impact ofNADPH engineering onGlaA production
isstrain‑dependent
All 14 strains were subjected to batch cultivations
in shake-flask format, whereby a medium contain-
ing maltose as GlaA-inducing carbon source was
used. FW35.1 (a pyrG + derivative of AB4.1) and B36
were taken along as corresponding reference strains.
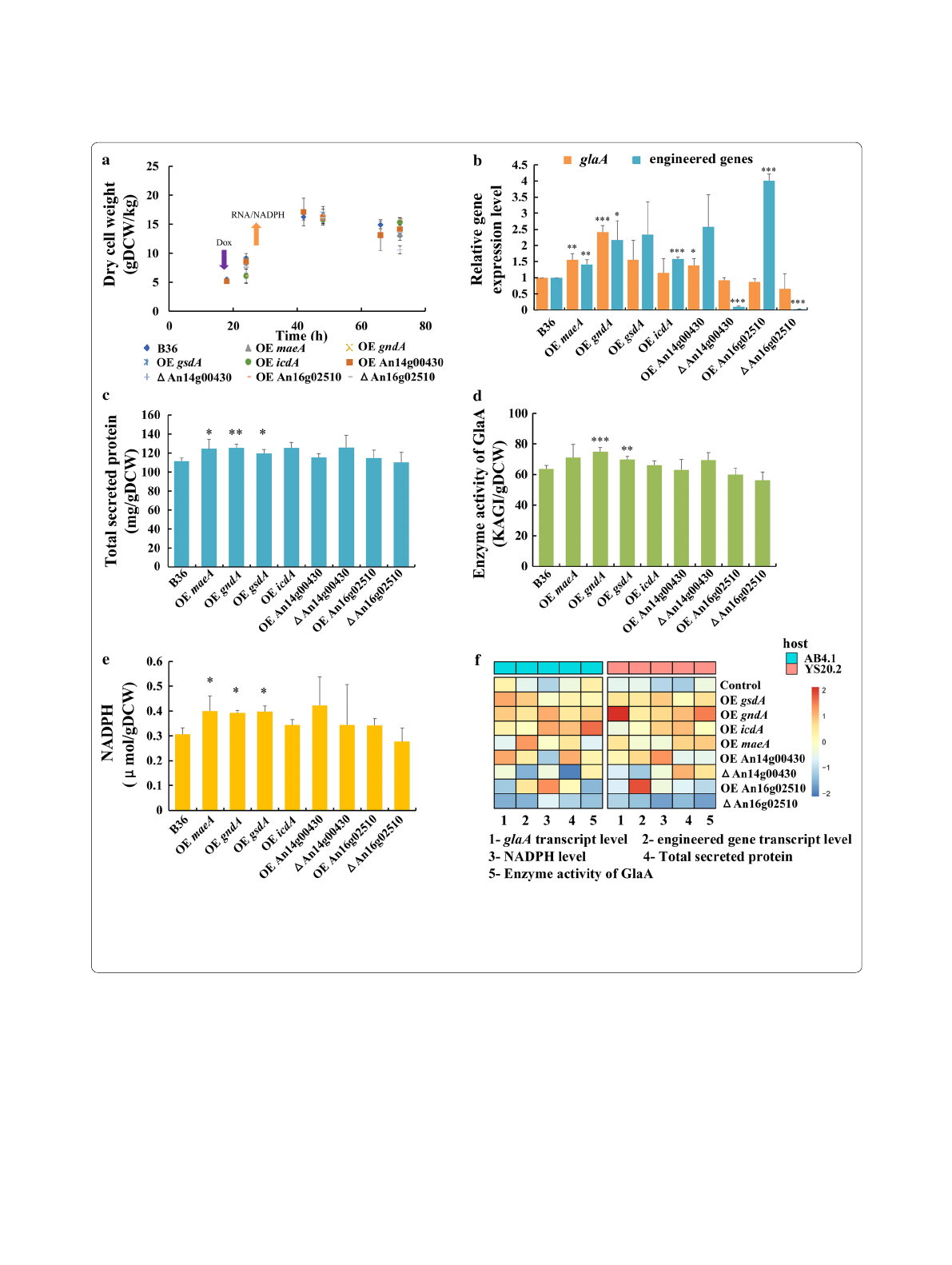
DOX-induced gene expression in all seven AB4.1
derivatives was about 1.5–2.7 times higher compared
to the reference strain FW35.1 as examined by qRT-
PCR (Additional file1: Fig. S8, TableS4). Although
this led to an elevated NADPH pool of about 30% in
Fig. 1 Pathway map highlighting all seven genes modified during this study in red. The cytosolic glycolytic pathway, the pentose phosphate
pathway and the mitochondrially located citric acid cycle are shown
Page 5 of 17
Suietal. Microb Cell Fact (2020) 19:198
the case of gndA, icdA or An16g02510, no significant
increase in GlaA enzyme activity was observed for all
of seven strains compared to the FW35.1 reference
(Additional file1: Fig. S8, TableS4). However, when
all seven candidate genes were overexpressed in the
YS20.2 background strain containing seven glaA gene
copies, increased transcript levels were similar as in
AB4.1, but for An16g02510 higher transcription levels
(fourfold) were observed (Fig.2b). Noteworthy, over-
expression of gndA displayed the highest effect on the
Fig. 2 Data for shake flask‑level cultivations of all engineered strains in the YS20.2 background in relation to the control strain B36. a Dry cell weight
(DCW); b Relative expression level of glaA and engineered genes; c Total secreted protein per gram biomass at 72 h after inoculation; d Enzyme
activity of GlaA per gram biomass at 72 h after inoculation; e Intracellular NADPH concentration in the exponential phase; f Comparison between
engineered strains in the AB4.1 and YS20.2 background, respectively. All experiments were conducted in biological quadruplicates. Significance
values were calculated with the two‑tailed t‑test with independent variables (*p < 0.05, **p < 0.01, ***p < 0.001)
Loading more pages...