
The Potential of Eukaryotic Cell-Free
Systems as a Rapid Response to Novel
Zoonotic Pathogens: Analysis of
SARS-CoV-2 Viral Proteins
Franziska Ramm
1
,
2
, Srujan K. Dondapati
1
, Hoai Anh Trinh
1
,
3
, Dana Wenzel
1
,
Ruben M. Walter
1
,
3
, Anne Zemella
1
and Stefan Kubick
1
,
2
,
4
*
1
Fraunhofer Institute for Cell Therapy and Immunology (IZI), Branch Bioanalytics and Bioprocesses (IZI-BB), Potsdam, Germany,
2
Institute of Chemistry and Biochemistry, Freie Universität Berlin, Berlin, Germany,
3
Department of Applied Biochemistry, Institute
of Biotechnology, Technical University Berlin, Berlin, Germany,
4
Faculty of Health Sciences, Joint Faculty of the Brandenburg
University of Technology Cottbus–Senftenberg, The Brandenburg Medical School Theodor Fontane, The University of Potsdam,
Potsdam, Germany
The ongoing pandemic caused by the novel coronavirus (SARS-CoV-2) has led to more
than 445 million infections and the underlying disease, COVID-19, resulted in more than 6
million deaths worldwide. The scientific world is already predicting future zoonotic
diseases. Hence, rapid response systems are needed to tackle future epidemics and
pandemics. Here, we present the use of eukaryotic cell-free systems for the rapid
response to novel zoonotic diseases represented by SARS-CoV-2. Non-structural,
structural and accessory proteins encoded by SARS-CoV-2 were synthesized by cell-
free protein synthesis in a fast and efficient manner. The inhibitory effect of the non-
structural protein 1 on protein synthesis could be shown in vitro. Structural proteins were
quantitatively detected by commercial antibodies, therefore facilitating cell-free systems for
the validation of available antibodies. The cytotoxic envelope protein was characterized in
electrophysiological planar lipid bilayer measurements. Hence, our study demonstrates
the potential of eukaryotic cell-free systems as a rapid response mechanism for the
synthesis, functional characterization and antibody validation against a viral pathogen.
Keywords: eukaryotic cell-free systems, SARS-CoV-2, viral proteins, rapid response, protein analytics, planar lipid
bilayer measurements
INTRODUCTION
Infectious diseases that are transferred from an animal to a human being, so-called zoonoses, can lead
to devastating health issues around the world as can be seen from the example of the severe acute
respiratory syndrome coronavirus type 2 (SARS-CoV-2). The close interaction with animals, such as
in agriculture and with domesticated animals (pets), the increasing consumption of different meats
as well as the intrusion of humans into the natural habitat of animals, causes a high risk for the
development of novel zoonoses that might lead to short lived disease outbreaks, epidemics or even
pandemics. Standard techniques such as the detection of viral antigens by polymerase chain reactions
(PCR) as well as rapid antigen tests accelerate prompt responses such as quarantines and shutting
down social contacts. Unfortunately, some pathogens, such as airborne viruses, are persistent and
have to be counteracted with vaccines and therapeutics. Consequently, the thorough characterization
of the virus itself and its mode of action, including the viral assembly, cell attack, pathogenesis of the
Edited by:
Simon J. Moore,
University of Kent, United Kingdom
Reviewed by:
Karen Marie Polizzi,
Imperial College London,
United Kingdom
Dong-Myung Kim,
Chungnam National University,
South Korea
*Correspondence:
Stefan Kubick
[email protected]aunhofer.de
Specialty section:
This article was submitted to
Synthetic Biology,
a section of the journal
Frontiers in Bioengineering and
Biotechnology
Received: 15 March 2022
Accepted: 01 April 2022
Published: 19 April 2022
Citation:
Ramm F, Dondapati SK, Trinh HA,
Wenzel D, Walter RM, Zemella A and
Kubick S (2022) The Potential of
Eukaryotic Cell-Free Systems as a
Rapid Response to Novel Zoonotic
Pathogens: Analysis of SARS-CoV-2
Viral Proteins.
Front. Bioeng. Biotechnol. 10:896751.
doi: 10.3389/fbioe.2022.896751
Frontiers in Bioengineering and Biotechnology | www.frontiersin.org April 2022 | Volume 10 | Article 8967511
ORIGINAL RESEARCH
published: 19 April 2022
doi: 10.3389/fbioe.2022.896751
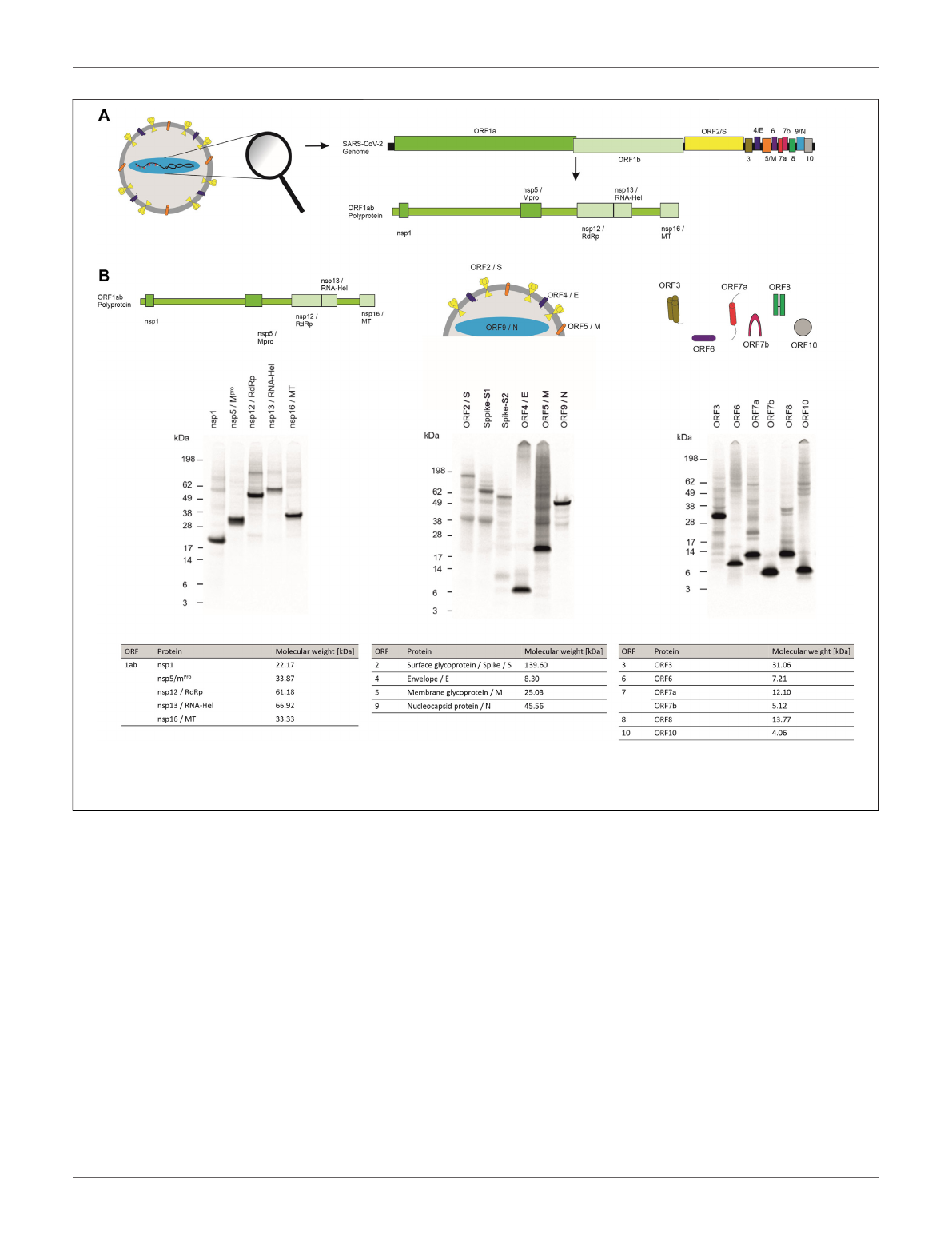
underlying disease and characterization of the viral proteins are
necessary to tackle these tasks and to identify efficient drugs. It is
essential to understand the different modes of action of the
individual proteins as each particular component plays a
specific role in the viral assembly, host infection and immune
invasion as well as viral replication. The novel coronavirus
expresses 10 different open reading frames (ORFs) encoding
the ORF1ab polyprotein, eight single ORFs corresponding to
eight single proteins as well as ORF7 which can be further
separated into ORF7a and b which encode two different
proteins (Figure 1A)(Kim et al., 2020;Wang et al., 2020;
Yoshimoto, 2020). The ORF1ab polyprotein encodes 16 non-
structural proteins (nsp) including proteases (nsp3 and nsp5), a
RNA-helicase (nsp13) and a RNA-dependent RNA-polymerase
(RdRp, nsp12) that are mainly responsible for viral replication
(Gao et al., 2020;Jang et al., 2020;Shin et al., 2020;Shu et al., 2020;
Yoshimoto, 2020;Zhang et al., 2020). This polyprotein is encoded
by the two ORFs ORF1a and ORF1b. A frameshift before the stop
codon in ORF1a facilitates the translation to be continued to
ORF1b and therefore resulting in the polyprotein ORF1ab (Kim
et al., 2020). The other ORFs can be divided into two major
classes, namely the structural proteins and the accessory proteins.
Structural proteins are well known and well characterized as these
proteins assemble to the viral capsid. The core structure of SARS-
CoV-2 virus is maintained by the ORF2 surface glycoprotein (S)
otherwise known as the Spike protein, the ORF4 envelope protein
(E), the ORF5 membrane glycoprotein (M) and the ORF9
nucleocapsid protein (N) (Figure 1A). The accessory proteins
of SARS-CoV-2 include membrane proteins like the putative ion
channel encoded by ORF3 (Kern et al., 2021), the type I
transmembrane protein coded by ORF7a (Rosenthal et al.,
2020) as well as the integral membrane protein ORF7b
FIGURE 1 | Cell-free synthesis of SARS-CoV-2 proteins in a CHO cell-free system. (A) Schematic figure showing the SARS-CoV-2 virus particle and its structure as
well as the coding sequences of the genome. (B) Qualitative analysis of cell-free synthesized SARS-CoV-2 proteins by autoradiography. Autoradiograph showing
14
C-leucine labeled proteins synthesized in a CHO system from the translation mixture (TM).
Frontiers in Bioengineering and Biotechnology | www.frontiersin.org April 2022 | Volume 10 | Article 8967512
Ramm et al. Analysis of SARS-CoV-2 Viral Proteins

(Schaecher et al., 2008). Accessory proteins ORF6 and ORF8 are
known to be involved in the host cell immune invasion and in
interferon signaling (Miorin et al., 2020;Flower et al., 2021;
Zhang et al., 2021). In some infections the protein encoded by
ORF10 is present whilst in others this protein cannot be found
which might be caused due to a read through of this ORF (Pancer
et al., 2020;Hassan et al., 2021). The proteins encoded by SARS-
CoV-2 and their characteristics are summarized in Table 1.
In order to characterize such a versatile set of proteins, a
system that can produce and characterize all kinds of proteins is
needed. Therefore, we present eukaryotic cell-free systems as a
promising methodology for the fast and efficient synthesis and
characterization of viral proteins based on the example of SARS-
CoV-2 proteins. In cell-free protein synthesis a crude cell lysate
rather than viable, intact cells are used, allowing the synthesis of
“difficult-to-express proteins”such as membrane proteins or
even cytotoxic proteins (Chalmeau et al., 2011;Orth et al.,
2011;Henrich et al., 2015;Thoring et al., 2017). With the
utilization of a cell lysate, the production of genetically
modified organisms becomes obsolete. As a result no high
laboratory safety standards are necessary for the cell-free
production of toxic and viral proteins. In order to avoid
cloning procedures for the generation of templates encoding
viral proteins, PCR templates can be used for cell-free
synthesis (Sawasaki et al., 2002). Thus, a fast screening of
different mutants is possible. Another advantage of some
eukaryotic cell-free systems are endogenous microsomal
vesicles derived from the endoplasmic reticulum (ER) that are
present in the lysate. These vesicles enable post-translational
modifications (PTMs) and are a natural surrounding for
membrane proteins (Brödel et al., 2014). Here, we
demonstrate that the whole set of viral proteins derived from
SARS-CoV-2 including membrane proteins, enzymes as well as
modulatory proteins can be synthesized in a lysate based on
Chinese hamster ovary (CHO) cells (Brödel et al., 2014;Thoring
et al., 2017). The synthesis of functional protein was verified by
cell-free synthesized nsp1 downregulating the in vitro synthesis of
a model protein and the demonstration of cytotoxic events as well
as single channel events in planar lipid bilayer measurements
induced by the cell-free synthesized envelope protein. Further,
the cell-free synthesized nucleocapsid protein was used to
demonstrate the possibility to quantitatively evaluate the
binding of commercially available antibodies. Taken together
eukaryotic cell-free systems, including but not limited to the
use of CHO lysate, can be applied to characterize viral proteins
and might facilitate the screening of antibodies as well as
pharmaceuticals and blockers against these viral proteins.
RESULTS
Viral pathogens such as SARS-CoV-2 induce cytotoxic effects
often associated with severe damage to the host cell. This might be
one of the major factors in the pathology and disease caused by
viruses. A valid system to characterize novel viral pathogens
should be able to synthesize and characterize structural as well
as non-structural proteins. Therefore, we used a eukaryotic cell-
free system to synthesize non-structural, structural and accessory
proteins encoded by SARS-CoV-2 (Figure 1A). Qualitative
analysis of viral proteins synthesized in a CHO cell-free
system showed that all viral proteins tested, could be
synthesized. Additionally, multimerization of proteins such as
ORF3 and ORF7a as well as ORF8 and ORF10 was visualized and
defined cleavage products as seen for Spike proteins and
nucleocapsid protein were detected by autoradiography
(Figure 1B).
Quantitative analysis by hot TCA precipitation and
subsequent liquid scintillation verified the acquired data for
the qualitative analysis. Total protein yields for non-structural
proteins showed that these proteins were mainly present in a
soluble form as higher protein yields were detected in the
supernatant fraction (SN) compared to the microsomal
fraction (MF). Cell-free protein synthesis of the full length
Spike protein (ORF2/S) was conducted using a PCR template
which resulted in a lower template concentration used. This
reduced template concentration led to lower total protein
yields. Nonetheless, the high molecular weight protein could
be synthesized in an equal amount to the comparably small
TABLE 1 | SARS-CoV-2 viral proteins and their characteristics.
ORF Protein Short Form Function
1ab Nsp1 —Inhibition of protein translation
Nsp5 m
Pro
Protease
Nsp12 RdRp RNA-dependent RNA polymerase
Nsp13 RNA-Hel RNA-helicase
Nsp16 MT Methyltransferase
2 Surface glycoprotein Spike/S Binding of host cells
3ORF3 —Ion channel
4 Envelope E Facilitates assembly
5 Membrane protein M Interacts with E, S and N to form stable assembly
6ORF6 —Immune invasion
7 ORF7a —Virus-host interaction, the type I transmembrane protein
ORF7b —Virus-host interaction, integral membrane protein
8ORF8 —Immune invasion
9 Nucleocapsid protein N Genome packaging
10 ORF10 —Unclear
Frontiers in Bioengineering and Biotechnology | www.frontiersin.org April 2022 | Volume 10 | Article 8967513
Ramm et al. Analysis of SARS-CoV-2 Viral Proteins
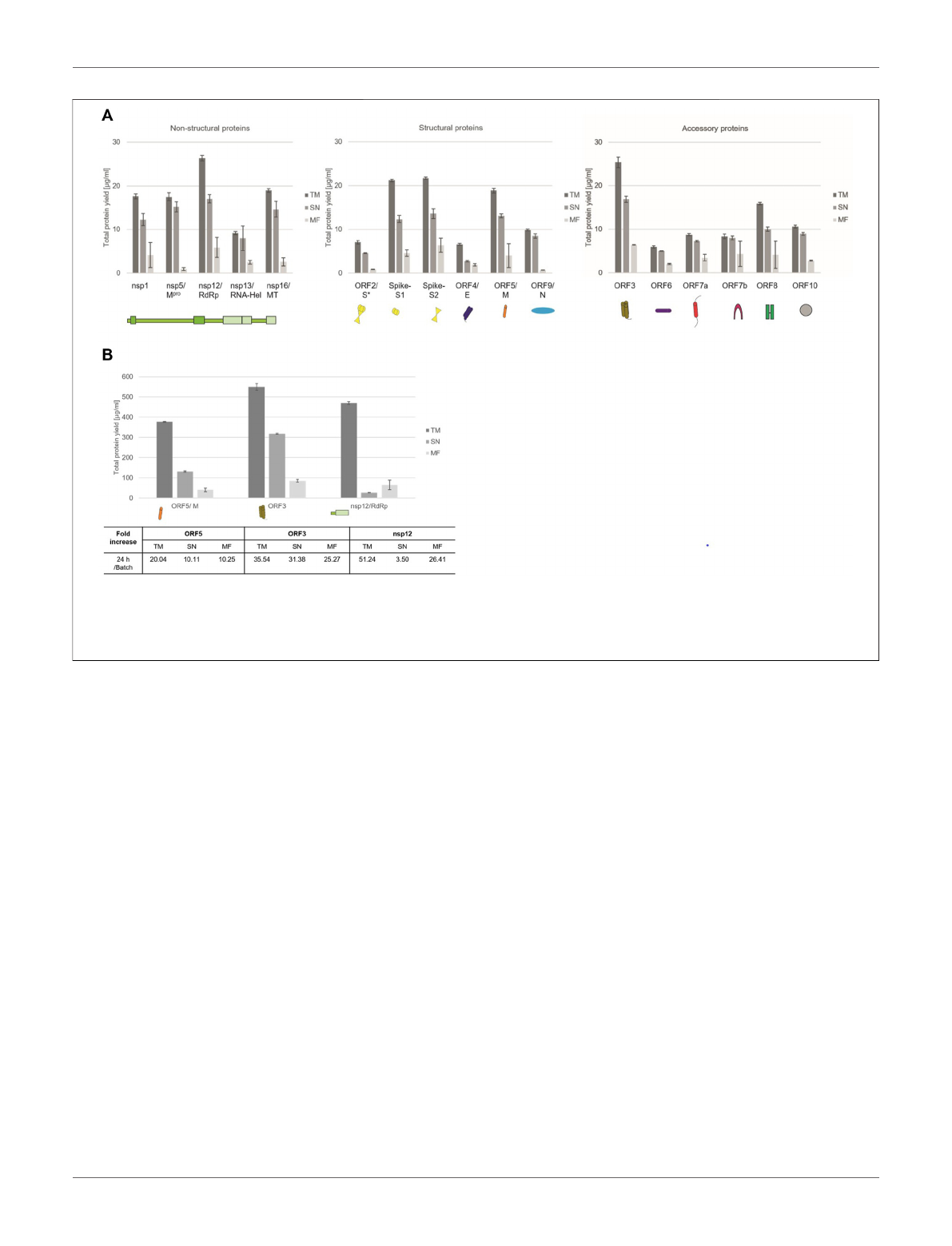
envelope protein (ORF4/E). The initial data for the synthesis of
the accessory proteins showed that all of these proteins could be
synthesized as well. The immunomodulatory protein encoded by
ORF6 showed the overall lowest protein yield of 6 μg/ml while the
transmembrane protein encoded by ORF3 showed the highest
protein yields with 25 μg/ml (Figure 2A).
In order to increase the protein yields in a continuous-
exchange cell-free (CECF) system, one representative protein
of the three protein groups was synthesized for 24 h. The
nsp12 coding for an RNA-dependent RNA polymerase
(RdRp), the ORF5 membrane glycoprotein and the channel-
like ORF3 were chosen (Figure 2B). The protein yields from a
batch-based synthesis could be increased by about 20, 35 and 50
fold for ORF5, ORF3 and nsp12, respectively, in a 24 h CECF
reaction. Apparently, the nsp12 enzyme was not suitable for a
CECF reaction as the soluble protein aggregated in the MF which
suggests that a batch-based reaction was more suitable for this
enzyme. These data indicate that CFPS offers a platform for the
rapid synthesis and analysis of SARS-CoV-2 proteins. As each
protein showed different requirements for the cell-free synthesis,
the open cell-free system offers an easy way to adapt the synthesis
conditions to the need of each individual protein.
To further show the applicability of CFPS as a rapid response
system for viral pathogens, we analyzed the individual protein
groups and analyzed the functionality of proteins of interest. In a
first step, cell-free synthesized nsp1 protein was characterized.
This protein is also known as the leader protein responsible for
the inhibition of host protein translation (Banerjee et al., 2020;
Thoms et al., 2020;Lapointe et al., 2021). It was further shown
that nsp1 did not decrease the translation of viral mRNA
(Banerjee et al., 2020), thus nsp1 was synthesized without any
alterations in the cell-free synthesis scheme. The nsp1 protein was
pre-synthesized in a cell-free manner and was added to the cell-
free synthesis of the model protein enhanced yellow fluorescent
protein (eYFP). Subsequently, the fluorescence intensity of eYFP
was measured during a 3 h synthesis time (schematic
representation in Figure 3A). The fluorescence signal of eYFP
without the supplementation of nsp1 was set as a baseline value of
100%. All other data were normalized to this intensity. In an
initial experiment pre-synthesized nsp1 from a CECF reaction, a
volume equivalent NTC and ORF6 as a protein control were
added to the eYFP synthesis. These data showed that the addition
of nsp1 at concentrations of 1000 and 600 nM reduced the
fluorescence intensity of eYFP in a concentration dependent
manner. 1000 nM of nsp1 led to a complete inhibition of
eYFP fluorescence. Unfortunately, the addition of the NTC
and ORF6 at 600 nM showed interactions with the eYFP
fluorescence. Nonetheless, 600 nM nsp1 induced the highest
FIGURE 2 | Quantitative analysis of cell-free synthesized SARS-CoV-2 proteins. Viral proteins were synthesized in CHO lysate in (A) batch-based reaction and (B) a
CECF reaction. The fold increase of total protein yield from a batch reaction to a CECF reaction is shown. Quantitative analysis of
14
C-labeled cell-free synthesized
proteins was performed by liquid scintillation counting. Standard deviations were calculated from triplicate analysis. The translation mixture (TM) was separated into the
soluble proteins in the supernatant (SN) and the microsomal fraction (MF). The template for the full length Spike protein was based on a PCR-template (*).
Frontiers in Bioengineering and Biotechnology | www.frontiersin.org April 2022 | Volume 10 | Article 8967514
Ramm et al. Analysis of SARS-CoV-2 Viral Proteins
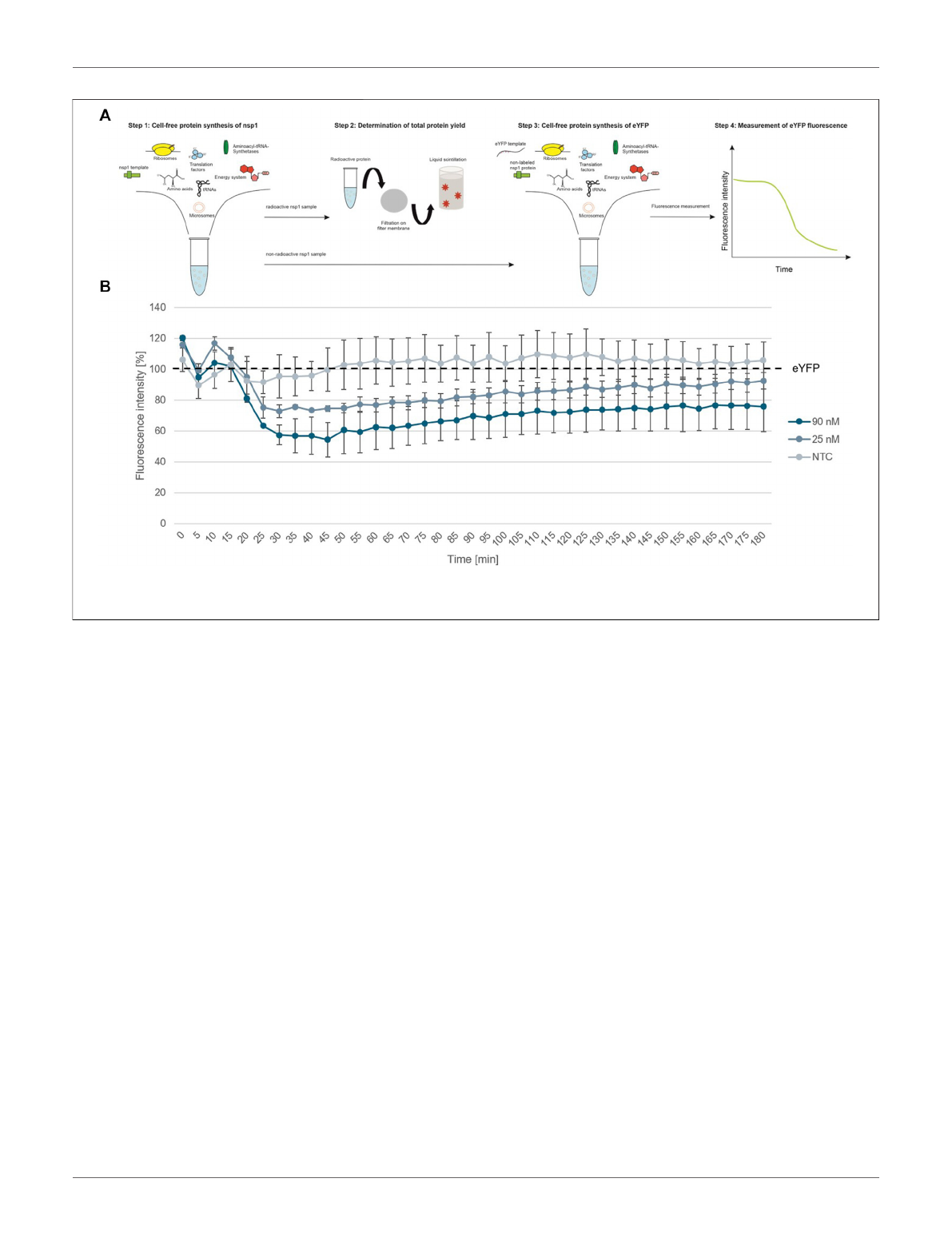
fluorescence decline (Supplementary Figure S1). Therefore, the
reaction conditions were optimized and a batch-based reaction
was used. Two concentrations (25 and 90 nM) of nsp1 protein
were added to the eYFP synthesis. The NTC was administered in
a volume equivalent to the highest nsp1 concentration. Starting
after about 20 min reaction time, the fluorescence intensity of the
eYFP slowly decreased when nsp1 was supplemented to the
reaction but no specific effect could be seen for an NTC
supplementation (Figure 3B). A concentration of 90 nM of
nsp1 decreased the eYFP fluorescence to less than 60%, while
25 nM nsp1 decreased the eYFP fluorescence to about 75%. This
suggested a concentration dependent effect. Strikingly, the
inhibitory effect of nsp1 weakened over time so that the
fluorescence intensity of eYFP increased again (Figure 3B).
Nonetheless, the translation inhibition effect of nsp1 could
be shown.
The characterization of non-structural viral proteins is
essential for a rapid response to a virus. Nonetheless, the virus
needs accessory proteins to stably infect the host. In SARS-CoV-2
these are known to trigger a variety of different interactions
within the host such as the involvement in the interferon
signaling pathway (Miorin et al., 2020;Yuen et al., 2020),
causing an immune invasion (Flower et al., 2021) or acting as
membranous channel-like proteins in order to disrupt the host`s
cell homeostasis (Yoshimoto, 2020;Kern et al., 2021). As shown
in Figure 1, all accessory proteins could be synthesized in a cell-
free manner and showed that cell-free protein synthesis can be a
tool to rapidly analyze the synthesis of such proteins, to identify
optimal synthesis conditions as well as to assess their solubility. In
the beginning of a pandemic, the role of such proteins is not yet
fully known, therefore we tested whether a signal peptide
interferes with a defined protein or might even inhibit the
translation of such a protein. Accordingly, ORF3, ORF6,
ORF7b, ORF8, and ORF10 were synthesized with and without
a Melittin (Mel) signal peptide, which typically allows for the co-
translational translocation. As some proteins might inhibit the
protein translation machinery, the Mel signal peptide allows for
the translocation of the protein into the vesicles present in the
cell-free lysates, enabling protein translation. All constructs with
and without a signal peptide were fractionated into the soluble
proteins (SN) and the proteins in the microsomal fraction (MF).
In general, our data indicated that all ORFs were more stably
expressed in the presence of a signal peptide (Figure 4A). The
NCM-ORF3 construct was based on a PCR template which might
have resulted in a lower overall translation efficiency due to a
lower template concentration. Strikingly, ORF10 protein
translation could not be quantitatively detected without a
signal peptide (Figure 4A, red box). In the presence of a Mel
signal peptide, protein yields of 13.7 μg/ml were detected for
ORF10 suggesting a better translation initiation. ORF7a is known
to harbor a native signal peptide as it encodes a type I
transmembrane protein (Rosenthal et al., 2020). Comparing
the native signal peptide (Nat-SP) with the Mel signal peptide,
the native signal peptide showed a higher total amount of protein
FIGURE 3 | Functional analysis of nsp1. (A) Schematic scheme of nsp1 functionality assay. (B) Application of cell-free synthesized nsp1 at 90 and 25 nM to the cell-
free protein synthesis of the model protein eYFP. Data were normalized to eYFP fluorescence signal. Standard deviation was calculated from two experiments.
Frontiers in Bioengineering and Biotechnology | www.frontiersin.org April 2022 | Volume 10 | Article 8967515
Ramm et al. Analysis of SARS-CoV-2 Viral Proteins
Loading more pages...